Clear Sky Science · zh
表观遗传记忆的随机建模
细胞如何记住它们的身份
你体内的每个细胞携带的基本上是相同的 DNA,但大脑细胞与皮肤细胞或免疫细胞的行为却截然不同。这种持久的“身份感”必须在细胞分裂时被复制,当复制失败时,诸如癌症的疾病可能发生。本文解释了研究者如何使用数学工具来理解表观遗传记忆——DNA 及其包装上的生化标记,使细胞能够长期记住某些基因应当保持开启还是关闭。
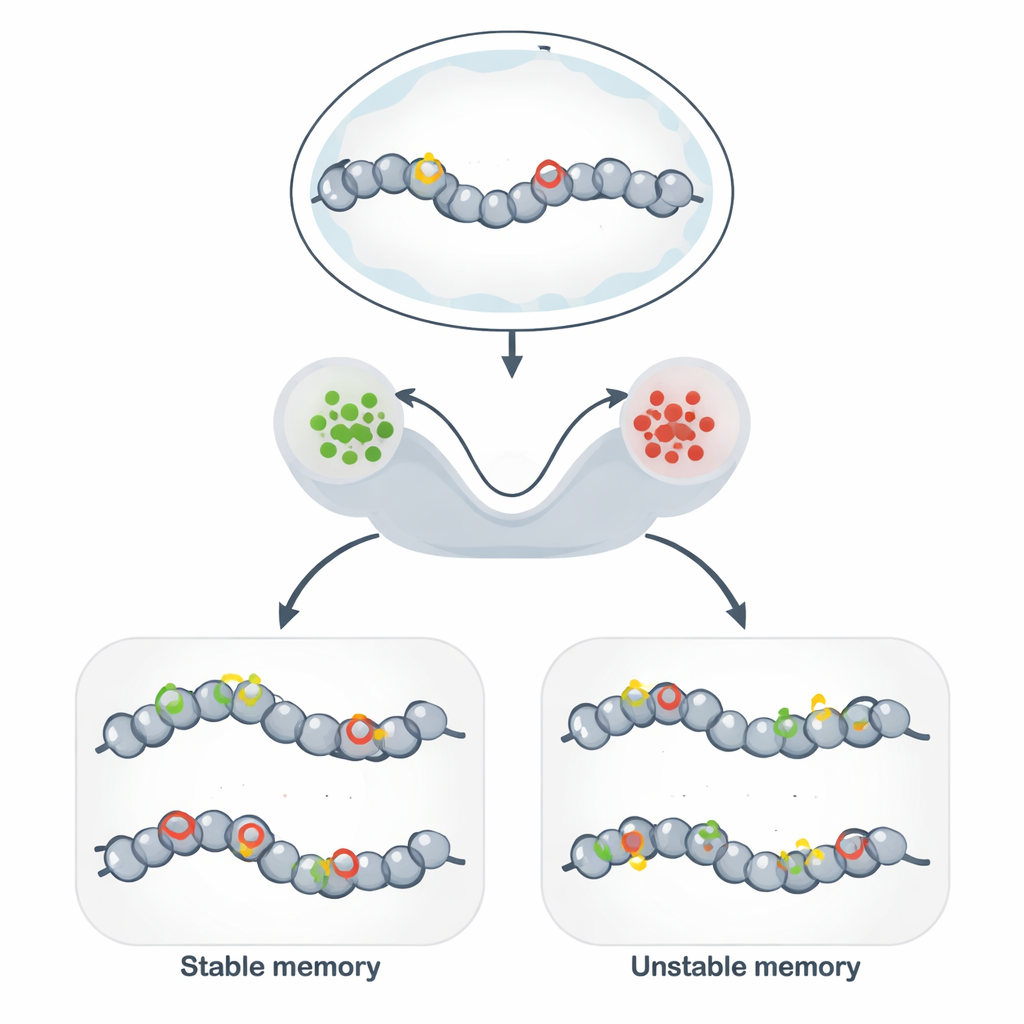
超越 DNA:基因组上的稳定标记
表观遗传记忆使得基因相同的细胞可以在许多细胞分裂中维持不同且稳定的行为。细胞并不改变 DNA 序列,而是在 DNA 及其周围的蛋白上装饰化学标签,使局部结构松弛或压缩。当标签有利于开放结构时,附近基因往往呈现活跃状态;当标签促成紧密结构时,基因则被沉默。这一机制构成了细胞类型身份、免疫应答的持久性、某些形式的大脑记忆,以及合成生物学中旨在“编程”稳定细胞状态的有前景工具的基础。
为何不能忽视随机性
在分子尺度上,细胞是一个嘈杂的环境:关键分子往往数量很少,彼此随机碰撞。传统以平滑曲线描述基因活动随时间变化的模型可以预测多重稳定态,类似两个由山峰分隔的山谷。在这些模型中,一旦系统滚入某个山谷就应永远停留。然而现实中,随机的分子事件有时会将系统从一边踢过山峰,进入另一山谷。本文介绍了将反应视为随机事件的随机(概率)框架,并量化此类开关发生的频率,例如通过平均首次通过时间来衡量在噪声触发跳转之前某一状态通常能维持多久。
观察分子混乱的数学透镜
综述描述了两种主要的随机建模方法。随机化学反应网络跟踪精确的分子计数,并将系统视为马尔可夫过程,其中每个反应以依赖当前状态的概率发生。这种方法能忠实捕捉稳定基因表达状态之间的罕见切换,但计算代价高。第二类模型使用随机微分方程,将分子计数近似为受高斯噪声扰动的连续量。化学朗之万方程和线性噪声近似等变体以牺牲部分精度换取解析清晰度,使研究者得以推导系统参数如何塑造稳定态附近的波动以及态间跃迁的可能性。
作为记忆硬件的染色质电路
为将这些抽象模型与生物学连接,作者们聚焦于染色质修饰电路:组蛋白和 DNA 上的化学标记网络相互加强。促进活性染色质的组蛋白标签和促进抑制的标签都能招募酶,将相同标签扩散到邻近位点,形成强大的正反馈。当这种自我强化比擦除过程快时,随机模型显示系统会发展出两种长期存在的状态——大致的“开启”和“关闭”——作为二元记忆。当加入在 DNA 复制期间被复制的高度稳定标签——DNA 甲基化时,系统更偏向长期抑制,并能显著延长沉默状态的持续时间。
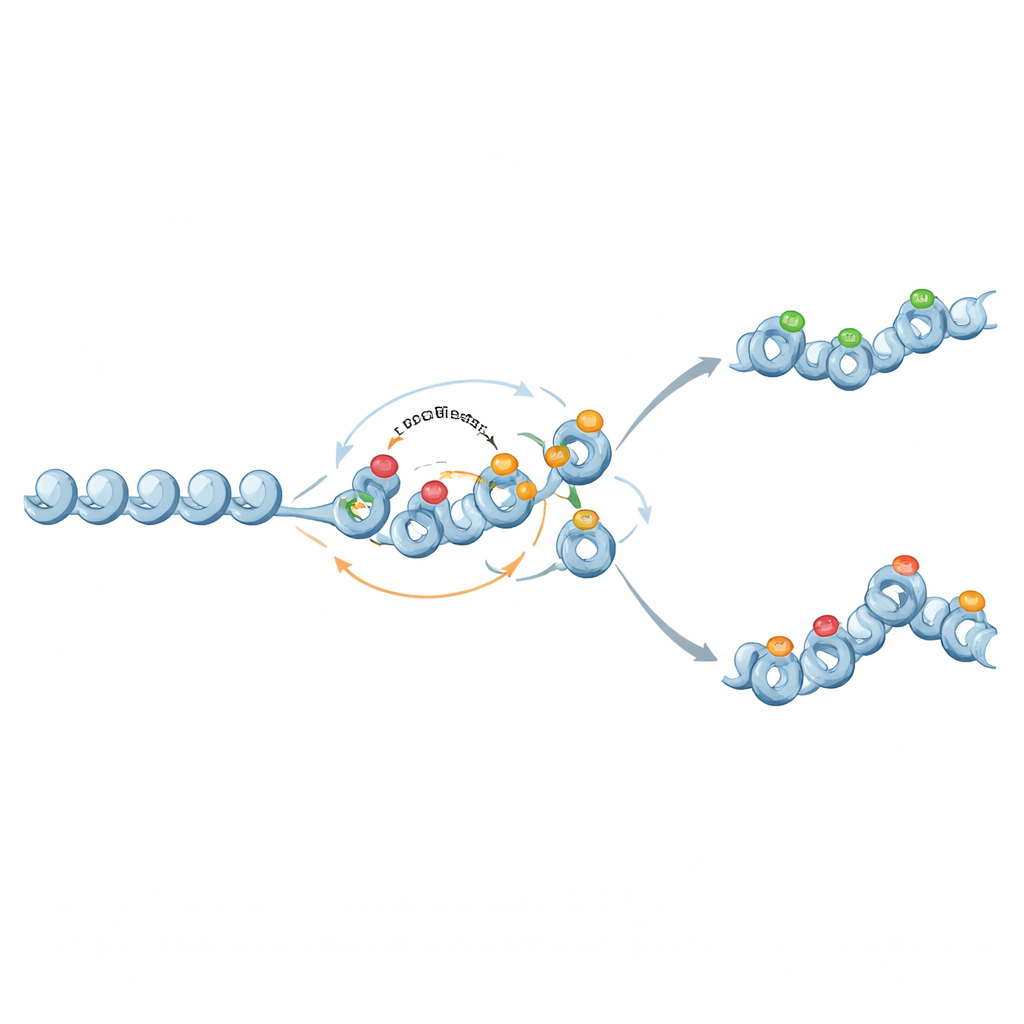
从二元开关到梯度记忆
有趣的是,相同的分子成分也能支持更微妙的类比记忆。当来自抑制性组蛋白标记到 DNA 甲基化的反馈减弱,且甲基化变得几乎永久但不具自我放大性时,甲基化充当一个固定的背景,设定基因表达的平均水平。组蛋白标记则在该背景上波动,有时在中等甲基化水平产生两种稳定的染色质状态和宽分布的表达。在这种方式下,染色质电路可以调节基因活动的稳定性以及细胞间表达的可变性。
这些模型为何对医学与设计重要
作者总结认为,理解表观遗传记忆需要明确建模分子噪声和关键过程的时间安排。他们的综述强调了哪些反应速率和反馈回路最强烈地控制细胞在活性或被抑制状态中停留的时间。这些见解可以指导改变细胞周期速度、酶水平或特定染色质标记的实验以检验模型预测。从长远来看,此类模型可能有助于设计可被可靠重编程、保持期望命运或以可控方式存储信息的细胞——为再生医学、癌症治疗和可编程免疫反应开辟了可能性。
引用: Bruno, S., Del Vecchio, D. Stochastic modeling of epigenetic memory. npj Syst Biol Appl 12, 58 (2026). https://doi.org/10.1038/s41540-026-00664-9
关键词: 表观遗传记忆, 染色质修饰, DNA 甲基化, 随机建模, 基因调控