Clear Sky Science · he
מודל חישובי מבוסס-כללים של ויסות מערכת MAPK/ERK
למה מתגים זעירים בתא חשובים
כל שנייה, התאים שלנו צריכים להחליט האם לגדול, להתחלק, לנוע או להתאבד במכוון. מקבל ההחלטות המרכזי הוא שרשרת חלבונים הנקראת מסלול MAPK/ERK. זהו גם אחד המערכות שנפגעות לעתים התכופות ביותר בסרטן. על אף שספרי לימוד מציגים את המסלול כרלאי פשוט, למעשה זהו רשת צפופה של רכיבים מצטלבים. המחקר הזה בונה מודל מחשב מפורט של אותה רשת כדי לשאול שאלה מעשית: כיצד מסלול אחד יכול בו-זמנית להנפיק תגובות מדודות ומדורגות וכן החלטות חדות של הכל-או-כלום, וכיצד חלבונים קרובים משתפים פעולה או מתחרים כדי לקבוע את גורל התא?
כביש מהיר עמוס בתוך התא
מסלול MAPK/ERK מתחיל במעטפת התא, שם גורמי גדילה נקשרים לקולטן ומגוייס חלבון מסייע בשם SOS. SOS מפעיל את RAS, מתג מולקולרי קטן המחובר לפנים פנימי של ממברנת התא. RAS פעיל מגייס משפחה של אנזימים הנקראים RAF, שמפעילים את MEK, אשר בתורו מפעיל את ERK. ERK מופעל ממשיך לווסת מאות מטרות בתא, ומעצב האם התא יתחלק, יתמיין, יגרום תנועה או יעבור מוות מתוכנת. המסלול, במקום להיות קו ישר פשוט, מלא בלופי משוב, הסתעפויות ווריאנטים מקבילים של אותם חלבונים (איזואפורמים), מה שהופך את התנהגותו לתלויית-הקשר במידה רבה.
ממפל פשוט לרשת מבוססת-כללים
כדי ללכוד את המורכבות הזו, המחברים משתמשים בדגום מבוסס-כללים, שיטה שמקודדת דפוסי אינטראקציה במקום לפרט כל מצב מולקולרי אפשרי בנפרד. המודל שלהם כולל שתי גרסאות של MEK (MEK1 ו-MEK2) ושלוש גרסאות של RAF (BRAF, CRAF ו-ARAF), וכן שותפים רגולטוריים כגון חלבוני 14-3-3. הוא בונה גם משוב מחזק אחד (שבו RAS פעיל מחזק את המפעיל שלו SOS) ושלושה משובי בלימה (שבו ERK פעיל מדכא את SOS, RAF ו-MEK). מכיוון שכל חלבון יכול לעבור שינויים ולהיקשר בדרכים מרובות, מודל מסורתי היה מתפוצץ בגודל; הגישה מבוססת-הכללים שומרת אותו ניהולית ועדיין מייצגת יותר מ-600 מינים מולקולריים מובחנים ואלפי תגובות.
הסבר לאותות מדרגתיים ולאותות דמויי-מתג
ניסויים הראו תבנית מבלבלת: ברמת RAS ו-RAF, הפעילות עולה בהדרגה ככל שמשתנה מינון גורם הגדילה, אך בתחתית המפל ERK לעתים נדירות נדלק באופן כמעט הכל-או-כלום. המודל מסביר זאת בכך שבמינונים נמוכים, RAS נדלק רק בכתמים קטנים של ממברנת התא. ככל שהמינון עולה, יותר חלקי ממברנה עוברים למצב פעיל. באופן מקומי, מתג RAS הוא דו-יציב — הוא קופץ בחדות מכבוי להדלקה — אך מכיוון שרק חלק מן הממברנה פעיל, הפעילות הממוצעת של RAS ו-RAF על פני כל התא נראית חלקה. במורד הזרם, ההפעלה הדו-שלבית של MEK ו-ERK פועלת כמגבר חזק, כך שגם פעילות מקדמית צנועה מייצרת הפעלה כמעט מלאה של ERK, ותוצאתה תגובה חדה בדמוי מתג. משוב שלילי מ-ERK ל-SOS ממיר אז גירוי מתמשך לפולסים קצובים של פעילות ERK, שתקופתם תלויה בעוצמת גורם הגדילה ובאינטנסיביות המשוב.
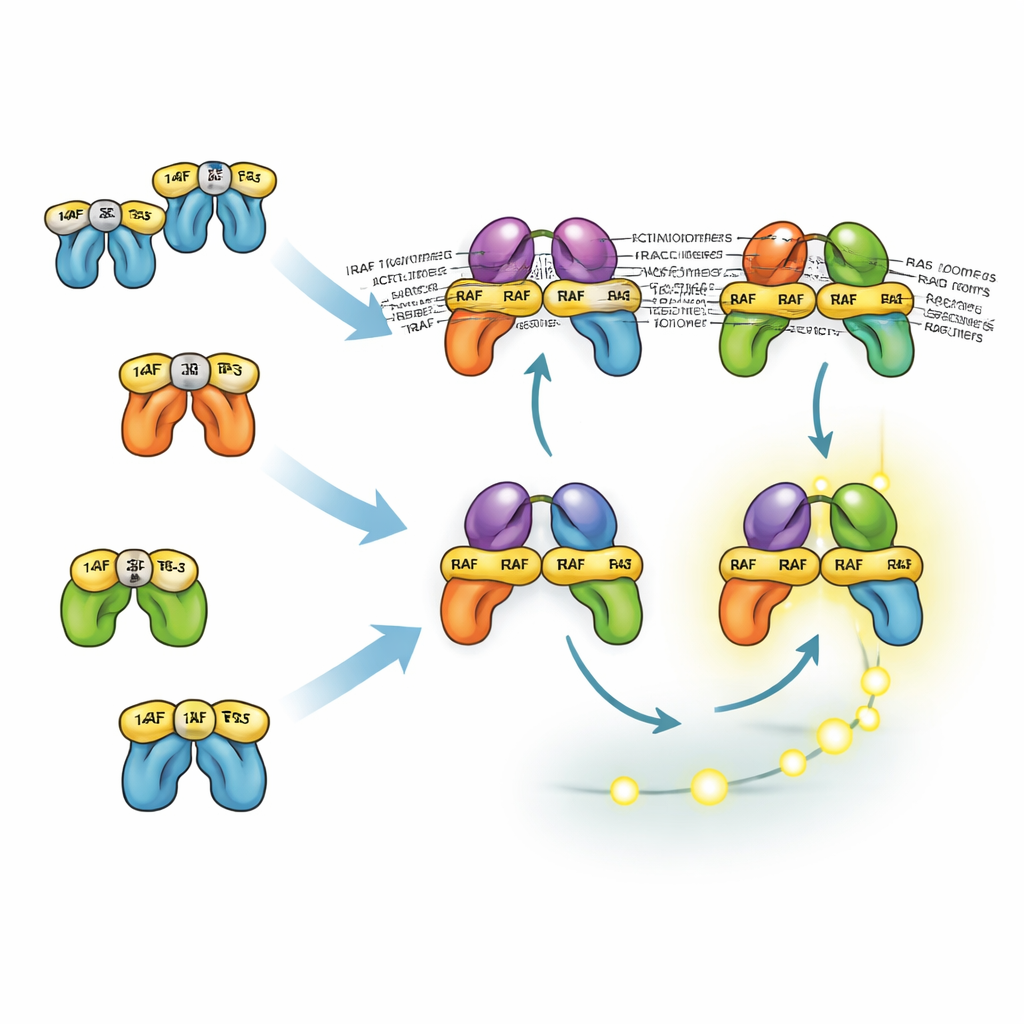
כיצד חלבונים דומים חולקים ומחלקים מטלות
שלושת האיזואפורמים של RAF מתנהגים אחרת בשל האינטראקציה שלהם עם חלבוני 14-3-3, שיכולים גם לנעול מולקולות RAF בצורה לא פעילה וגם לייצב זוגות RAF פעילים. אתר העיגון העיקרי של 14-3-3 ב-BRAF נמצא בזנבו, מה שמאפשר לו להישאר קשור ל-14-3-3 בעודו עדיין נקשר ל-RAS ומייצר דימרים עם CRAF או ARAF. דימרים מעורבים אלה יציבים ויעילים במיוחד בהפעלת MEK. לעומת זאת, CRAF ו-ARAF מסתמכים יותר על קשירה בקצה הראש, מה שחוסם את גיוסם ל-RAS. כתוצאה מכך, דימרים שחסרים BRAF פחות יציבים ובדרך כלל פחות נפוצים. עם זאת, המודל מראה שגם בלעדיו ניתן לתאים לייצר אותות ERK חזקים כי משוב חיובי מעלה וייצור שבתחתית מפצה. זה מסייע להסביר מדוע תרופות שמכוונות רק ל-BRAF לעתים קרובות מדכאות את ERK חלקית ונדרשות בשילוב עם מעכבי MEK או ERK.
לאזן תנועה, הישרדות ומוות
המודל עוקב גם אחרי תפקידים של RAF שמעבר להפעלת ERK. כאשר דימרים מתפרקים תחת משוב שלילי מונהג על-ידי ERK, צורות פתוחות של CRAF יכולות להיקשר לשותפים ששולטים בתנועת התא ובהישרדותו, כגון קינאזת ROKα או חלבון קידום-המוות MST2. ככל שרמות גורם הגדילה ושפע 14-3-3 משתנות, האיזון נוטה בין דימרים עשירים ב-BRAF המניעים ERK בעוצמה, קומפלקסים מבוססי CRAF ו-ARAF שמודלים תנועתיות, ובריכות שמשחררות את MST2 כדי לקדם מוות תאי. מפני שאיזונים אלו מגיבים בצורה חלקה לעוצמת הגירוי בעוד הפלט של ERK יכול להיות כמעט הכל-או-כלום, אותו מסלול יכול לכוונן היבטים עדינים של התנהגות ועדיין לקבל החלטות גורל מכריעות.
מה משמעות הדבר להבנת תאים
על ידי אריגה יחד של אינטראקציות ספציפיות איזואפורם, מתג מקומי בממברנה ולולאות משוב מקוננות, המודל מראה כיצד מסלול מולקולרי אחד יכול לשמש גם ככמתר וגם כמתג דיגיטלי. צירופים שונים בכמויות BRAF, CRAF, ARAF, MEK1, MEK2 ו-14-3-3, כפי שנצפו ברקמות וגידולים שונים, מובילים באופן טבעי לדפוסים דינמיים שונים של איתות ERK — מפולסים ועד נמלים ממושכים. זה מציע כי חיווט מותאם-תא של אותם רכיבים מרכזיים יכול להקצות מחדש את מסלול MAPK/ERK לתפקידים פיזיולוגיים מגוונים, וכי טיפולים יעילים יזדקקו להתחשב לא רק במטרות בודדות אלא בהקשר הרגולוטורי המלא שבו פועלים מתגים מולקולריים אלו.
ציטוט: Kocieniewski, P., Lipniacki, T. A computational rule-based model of MAPK/ERK system regulation. Sci Rep 16, 14437 (2026). https://doi.org/10.1038/s41598-026-44353-3
מילות מפתח: אותות MAPK ERK, איזואפורמים של RAF, ויסות 14-3-3, מודל מסלול חישובי, דינמיקת גורל התא