Clear Sky Science · zh
通过与噬菌体超分子组装触发的细菌免疫激活
细菌如何智胜其病毒入侵者
细菌不断受到称为噬菌体的病毒捕食,这些病毒可以在几分钟内将繁荣的微生物群落变成一片死寂。研究揭示了一种令人意外的新型细菌反击方式:构建巨大的分子环来感知到来的噬菌体,然后有意从内部关闭被感染的细胞。该工作不仅揭示了一种巧妙的细菌自我防御形式,还与我们自身先天免疫系统使用的策略相呼应,提示了生命在检测和阻止感染方面存在的深层共同主题。
一个具有双重身份的小小守护者
论文关注一种作者将其重命名为 RAZR 的细菌蛋白,该蛋白存在于大肠杆菌(Escherichia coli)和许多其他物种中。单独存在时,RAZR 无害:细胞可以大量产生它而不出现不良影响。然而当某些噬菌体感染时,RAZR 会突然变得致命——对宿主细胞以及病毒本身都是如此。该系统通过“中止感染”发挥作用:一旦 RAZR 被激活,被感染的细菌停止生长并有效地自我牺牲,从而阻止病毒复制并在群体中传播。理解 RAZR 如何从静默旁观者切换为细胞执行者,是这项研究的核心问题。 
翻转开关的病毒环
研究人员发现 RAZR 是被若干不相关噬菌体产生的特定蛋白激活的。在一种噬菌体中,一种名为 Gp77 的蛋白——很可能参与修复或使病毒基因组环化——充当了触发子。在其他噬菌体中,触发子是位于病毒衣壳关键通道处并帮助将 DNA 注入宿主的“门”蛋白。尽管这些蛋白作为单体在序列和结构上差异很大,但它们都有一个显著共同特征:它们组装成直径几乎相同的大环。通过冷冻电子显微镜,团队展示了 Gp77 形成 24 聚体的环,而门蛋白形成 12 聚体的环。RAZR 通常形成松散的线性链,当它缠绕这些病毒环时,会构建出一个巨大的、多层的圆形复合体,直径超过 25 纳米。
构建分子末日装置
在该复合体内,RAZR 汇聚了两个功能区:一个束缚病毒环的锌指“传感器”和一个切割 RNA 的 HEPN“刀片”。来自相邻 RAZR 分子的两个锌指片段结合每个病毒蛋白单元,确保 RAZR 以精确且高度协同性的方式覆盖环。这个环状支架将 RAZR 的 RNA 切割结构域强行排列成弯曲且紧密堆积的构型,使许多切割位点像圆锯的齿一样排列在外缘。破坏 RAZR 与噬菌体蛋白之间接触面的突变,或削弱 RAZR 分子相互黏附方式的改变,会削弱这一防御。一段连接感知与切割部分的小柔性连杆也被证明至关重要,帮助蛋白在贴合病毒环时采用正确的几何构型。
切碎 RNA 以阻止病毒扩散
一旦在噬菌体构建的支架上组装完毕,RAZR 就变成了强大的 RNA 破坏机器。作者表明被激活的 RAZR 不接触 DNA,但会切割多种单链 RNA:转运 RNA、核糖体 RNA,以及来自细菌和感染噬菌体的信使 RNA。在活细胞中,这种广泛的 RNA 裂解迅速阻断蛋白质合成,同时基本不影响 DNA 复制和转录。在体外实验中,将纯化的 RAZR 与其病毒触发子混合即可完全停止蛋白质合成。通过以这种粗放且非特异的方式终止翻译,该系统注定了被感染的细胞,同时也剥夺了噬菌体复制所需的分子工具。 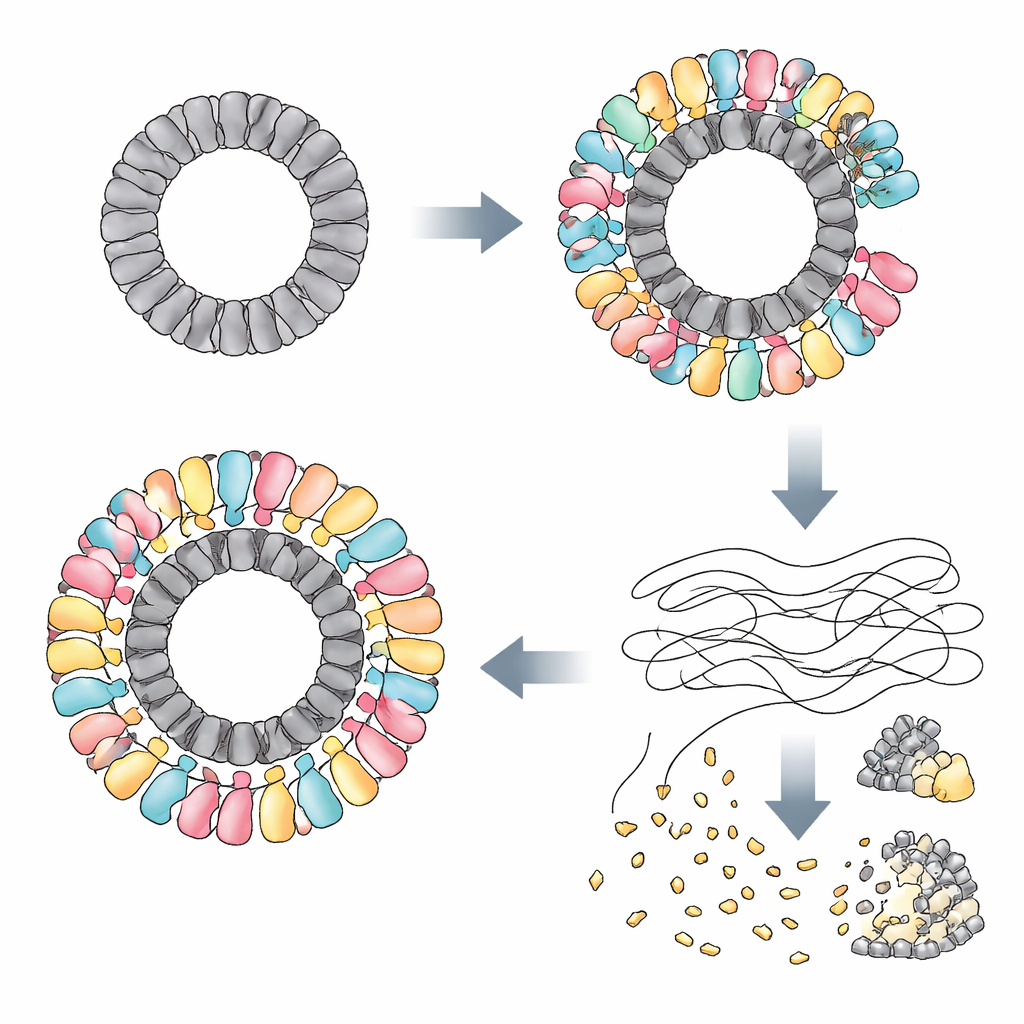
为不同病毒量身定制防御
尽管不同细菌来源的 RAZR 蛋白共享相似的切割结构域,研究发现它们的锌指传感器差异很大。将来自大肠杆菌和变异克雷伯菌(Klebsiella variicola)的两个 RAZR 版本之间互换这些传感器片段,会在很大程度上互换它们能阻止的噬菌体谱。即便两个 RAZR 变体针对相同噬菌体,它们也依赖病毒门蛋白上的略微不同接触点,这可从不同的病毒逃逸突变中看出。这表明细菌可以通过进化 RAZR 的传感器部分来微调其抗病毒范围,同时保留强大的 RNA 切割机器不变。
生命树上共享的理念
最后,作者提出 RAZR 感知的不是特定的病毒“密码”,而是一种结构性特征:在噬菌体感染期间出现、宿主自身机器中不存在的特定大小的大蛋白环。通过包裹这些环并将其用作自身组装的模板,RAZR 将微弱的病毒信号放大为全有或全无的免疫反应。这一策略与动物(包括人类)中许多先天免疫复合体形成巨大的丝状或环状结构以检测病原并激活破坏性酶的方式极为相似。因此该工作揭示了一个显著的趋同:从细菌到哺乳动物,生命不断演化出构建巨型蛋白结构的方法,将入侵病毒的几何形态反过来作为对抗它们的武器。
引用: Zhang, T., Lyu, Y., Beck, C.R. et al. Bacterial immune activation via supramolecular assembly with phage triggers. Nature 651, 1051–1059 (2026). https://doi.org/10.1038/s41586-025-10060-8
关键词: 噬菌体免疫, 细菌先天防御, 超分子蛋白复合体, RNA 裂解, RAZR 系统