Clear Sky Science · zh
通过多样结合途径动态控制IDP相互作用网络
细胞如何快速做出决策
在细胞内部,蛋白质不断在分秒之间选择配对、建立或断开连接。这些选择可以决定细胞是生长、修复损伤还是走向自毁。本文研究了来自抑癌蛋白p53的一个特别重要的片段如何在两个对立的配体之间快速切换:一个关闭p53活性,另一个则有助于激活它。通过观察单个分子的动态,作者表明这种切换并不沿单一路径进行,而是通过两条不同的途径,从而为细胞在应对应激时提供了更快且更灵活的反应方式。
会变形的蛋白枢纽
许多细胞内蛋白含有柔软、无固定结构的区域,称为本征无序蛋白(IDP)。与刚性、折叠良好的蛋白不同,IDP能够塑形以适配多种配体,在拥挤的分子网络中充当枢纽。研究组聚焦于p53起始端的转录激活域(TAD),这是一个典型的IDP片段,利用两个小区域(称为AD1和AD2)与众多调节因子相互作用。一个配体Mdm2促使p53降解并在细胞健康时维持其低水平;另一个配体Taz2属于一个大型共激活复合体,帮助p53启动应激反应基因。因为Taz2可以同时与AD1和AD2结合,而Mdm2仅结合AD1,p53 TAD成为研究单一柔性蛋白片段如何在具有相反影响的配体之间切换的理想系统。
观看单分子如何换配体
为了追踪这些快速的配体置换,研究者使用了三色单分子FRET,这是一种测量蛋白内部及其之间微小距离变化的荧光方法。他们在p53 TAD的端点和Taz2上分别连接了不同颜色的示踪染料,同时让Mdm2不被标记。通过将TAD固定在玻璃表面并以可控浓度加入配体,他们得以观察单个p53分子在结合、释放和重新结合Taz2与Mdm2时的动态。荧光模式的变化揭示了TAD是处于未结合状态、仅与Taz2结合、仅与Mdm2结合,还是短暂同时与两者结合形成三蛋白(三级)复合体的状态。
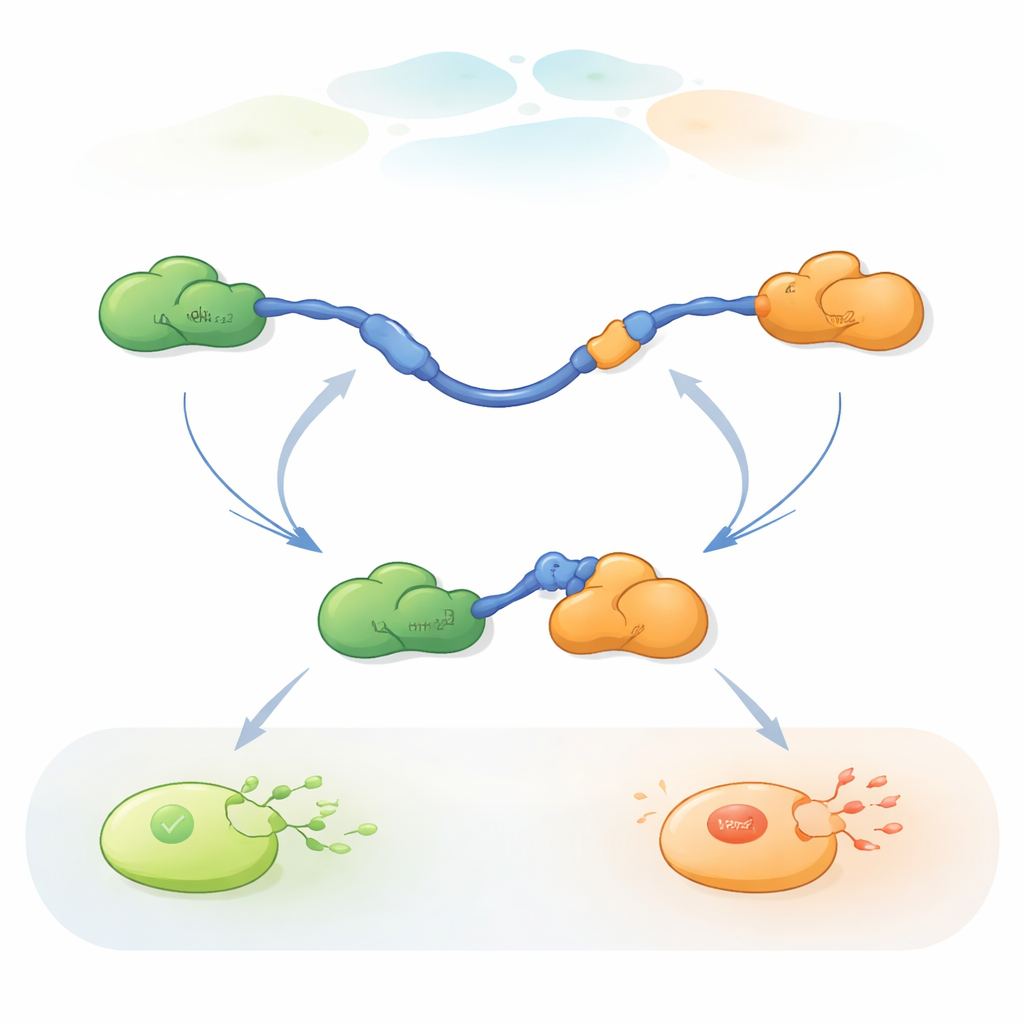
交换配体的两种途径
记录结果显示,TAD可以通过两种本质不同的方式交换配体。在竞争途径中,TAD先完全释放一个配体,变得自由且柔性,随后才与另一个配体结合。在变构途径中,TAD并不完全脱落,而是形成一个短寿命的三级复合体,Taz2与Mdm2同时结合,各自抓住TAD的不同部分。从这个联合状态出发,一个配体离开而另一个保持结合。通过统计TAD在每种状态下停留的时间及转换发生的频率,作者能提取出每一步的速率常数,并定量在不同条件下每条途径被使用的频率。
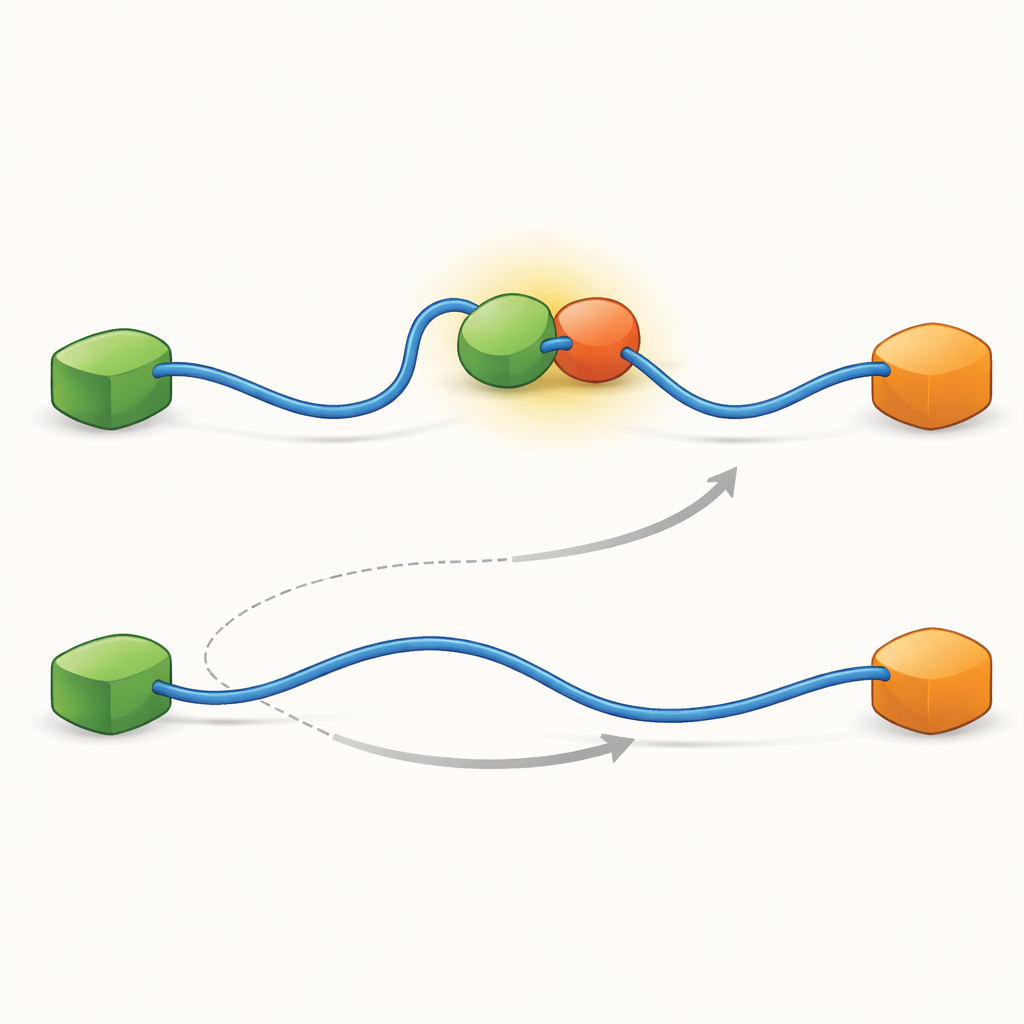
隐藏的中间步骤塑造网络行为
为了揭示使这两条途径得以存在的微观原因,团队将技术推向非常高的时间分辨率,并逐次分析光子到达事件。他们聚焦于TAD与Taz2结合的过程,发现该过程本身遵循两类不同的“转换路径”——即由未结合到完全结合之间的短暂中间阶段。在一种类型中,Taz2强烈抓住AD2而AD1保持游离;在另一种类型中,Taz2更多地作用于AD1而AD2相对松弛。AD2结合的中间体自然允许Mdm2滑上AD1并形成三级复合体,从而助长了变构途径。相反,AD1结合的中间体阻止了Mdm2结合,迫使系统走竞争路线,即TAD必须完全脱落后Mdm2才能结合。因此,TAD在结合过程中采用的微观、短暂构象最终决定了哪种大尺度的交换途径占主导。
关乎生死选择的快速切换
综合所有数据,作者表明三级复合体的变构途径可以承担Mdm2结合态与Taz2结合态之间的大部分流量,尤其是在细胞中Mdm2丰度较高的情况下。这一路径允许p53在条件改变时快速从被Mdm2抑制转为被Taz2激活,同时仍然限制过度的Mdm2重新结合。对TAD施加化学修饰以削弱Mdm2结合可将平衡重新推向竞争途径,提供了另一层调控。简而言之,该研究揭示了一个柔性蛋白片段利用其会变形的特性构建一个短暂的三方“握手”,以加快用一个配体换另一个配体的速度。这一机制可能是无序蛋白区域管理复杂信号网络并帮助细胞快速响应应激的普遍方式。
引用: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
关键词: 本征无序蛋白, p53调控, 蛋白质结合网络, 变构开关, 单分子FRET