Clear Sky Science · es
Control dinámico de la red de interacciones de IDP mediante vías de unión diversas
Cómo las células toman decisiones rápidas
En el interior de nuestras células, las proteínas están constantemente eligiendo compañeras, formando y rompiendo conexiones en fracciones de segundo. Estas decisiones pueden determinar si una célula crece, repara daños o se autodestruye. Este estudio examina cómo un segmento proteico especialmente importante del supresor tumoral p53 cambia rápidamente entre dos compañeras opuestas: una que apaga la actividad de p53 y otra que ayuda a activarla. Al observar moléculas individuales en acción, los autores muestran que este intercambio no sigue una única ruta simple, sino que utiliza dos vías distintas que proporcionan a la célula una forma más rápida y flexible de responder al estrés.
Un centro proteico que cambia de forma
Muchas proteínas en nuestras células tienen regiones flexibles y desestructuradas conocidas como proteínas intrínsecamente desordenadas (IDP). A diferencia de las proteínas rígidas y bien plegadas, las IDP pueden adaptarse para encajar con muchos socios distintos, actuando como centros en redes moleculares densas. El equipo se centró en el dominio de transactivación (TAD) al inicio de p53, un segmento IDP clásico que utiliza dos regiones pequeñas, llamadas AD1 y AD2, para interaccionar con numerosos reguladores. Un socio, Mdm2, promueve la degradación de p53 y mantiene sus niveles bajos cuando la célula está sana. Otro socio, Taz2, forma parte de un gran complejo coactivador que ayuda a p53 a activar genes de respuesta al estrés. Dado que Taz2 puede unirse tanto a AD1 como a AD2, mientras que Mdm2 se une solo a AD1, el TAD de p53 es un sistema ideal para preguntar cómo una sola pieza flexible de proteína puede compatibilizar compañeras con efectos opuestos.
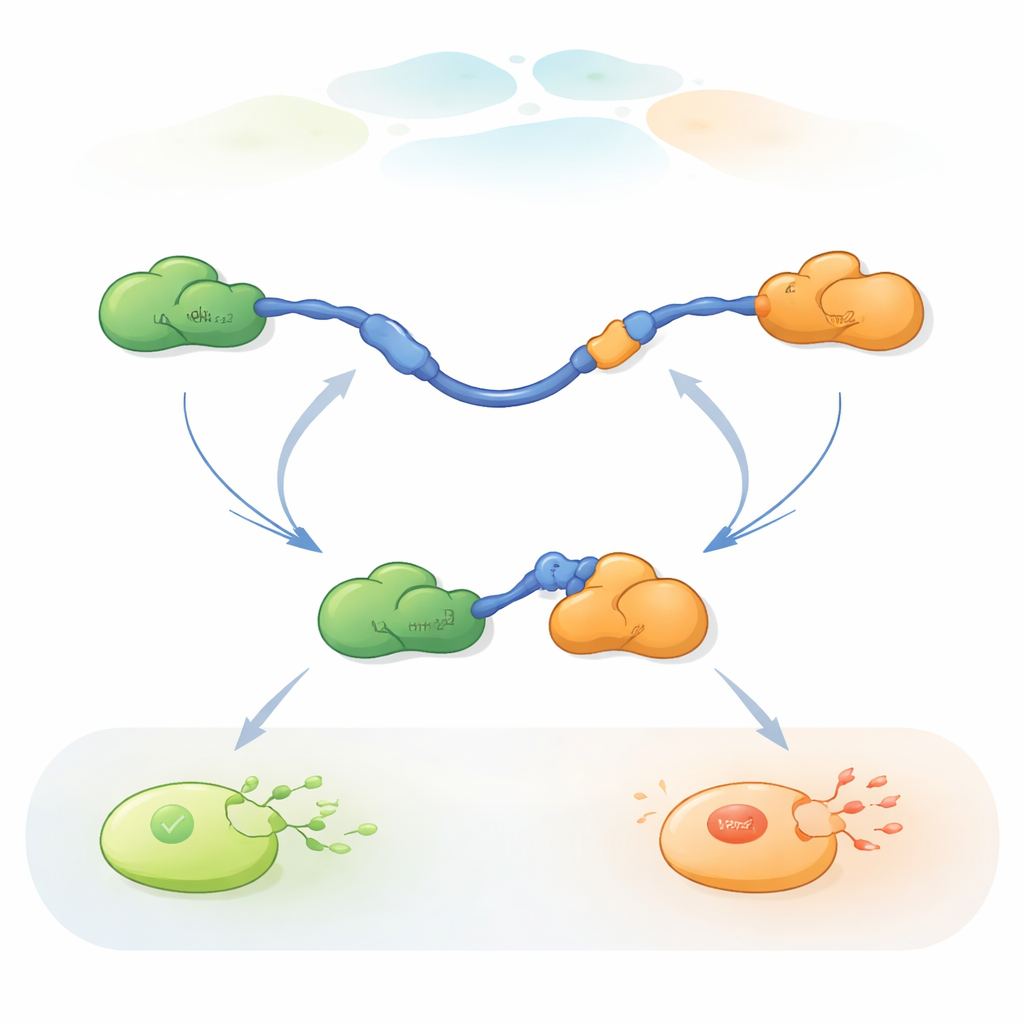
Observando moléculas individuales cambiar de compañera
Para seguir estos rápidos intercambios de compañeras, los investigadores utilizaron FRET tricolor de una sola molécula, un método de fluorescencia que mide cambios de distancia diminutos dentro y entre proteínas. Colocaron colorantes de distinto color en los extremos del TAD de p53 y en Taz2, mientras dejaban Mdm2 sin marcar. Al inmovilizar el TAD sobre una superficie de vidrio y añadir los socios en cantidades controladas, pudieron observar moléculas individuales de p53 mientras se unían, se liberaban y volvían a unirse a Taz2 y Mdm2 a lo largo del tiempo. Los cambios en el patrón de fluorescencia revelaron si el TAD estaba sin unión, unido solo a Taz2, unido solo a Mdm2, o brevemente unido a ambos a la vez en un complejo ternario de tres proteínas.
Dos maneras de intercambiar compañeras
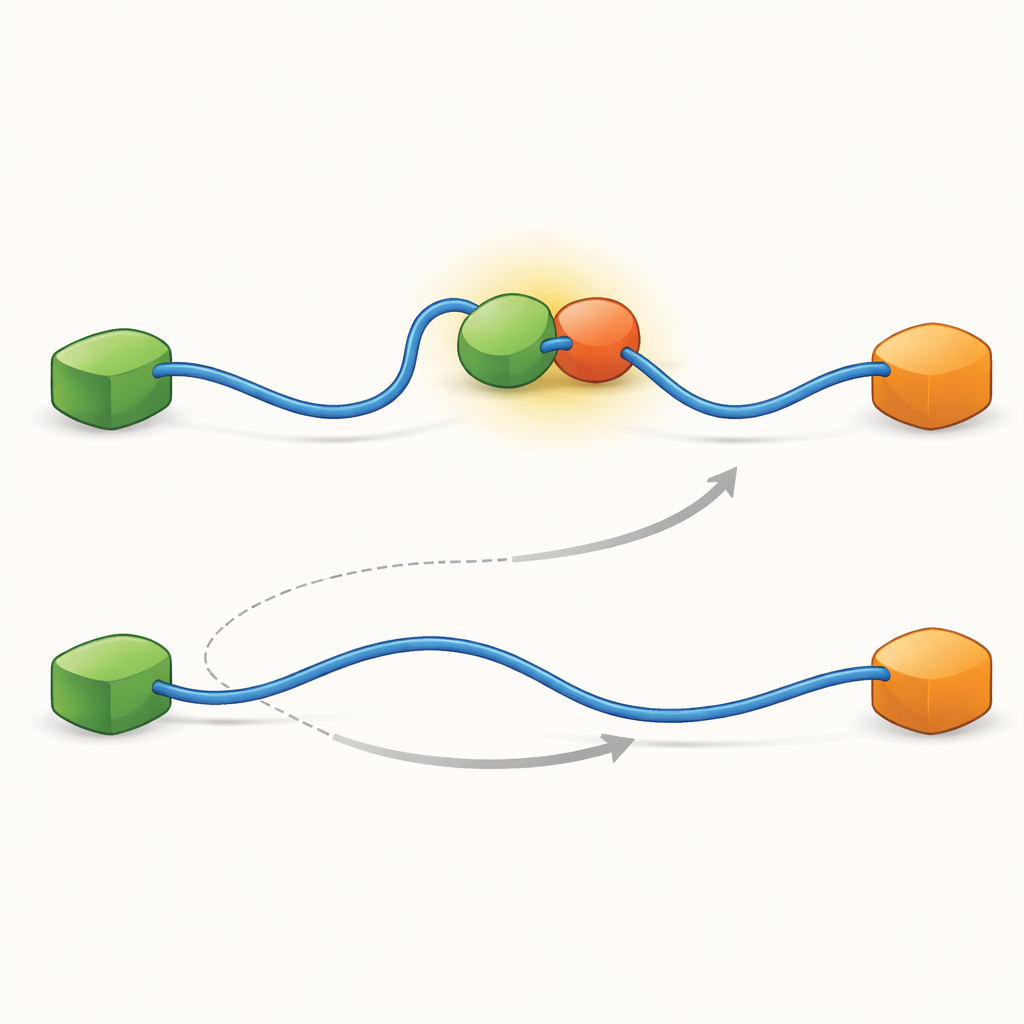
Las grabaciones mostraron que el TAD puede intercambiar compañeras de dos maneras cualitativamente diferentes. En la vía competitiva, el TAD primero suelta por completo a una compañera, quedando libre y flexible, y solo entonces se une a la otra compañera. En la vía alostérica, el TAD no se despega totalmente. En lugar de ello, forma un complejo ternario de corta duración en el que Taz2 y Mdm2 están ligados al mismo tiempo, cada uno agarrando una parte distinta del TAD. Desde este estado conjunto, una compañera se va y la otra permanece. Al contar cuánto tiempo el TAD permanecía en cada estado y con qué frecuencia ocurrían las transiciones, los autores pudieron extraer constantes de velocidad para cada paso y cuantificar con qué frecuencia se usa cada vía en diferentes condiciones.
Pasos intermedios ocultos modelan la red
Para descubrir qué hace posibles estas dos vías, el equipo llevó su técnica a una resolución temporal muy alta y analizó los eventos de llegada de fotones uno por uno. Se centraron en el acto de unión del TAD a Taz2 y encontraron que este proceso en sí sigue dos tipos distintos de “rutas de transición”: etapas intermedias breves entre los estados no unido y totalmente unido. En un tipo, Taz2 sujeta con fuerza a AD2 mientras AD1 queda libre; en el otro, Taz2 se compromete más con AD1 mientras AD2 está más flojo. El intermedio en el que AD2 está unido permite naturalmente que Mdm2 se coloque sobre AD1 y cree el complejo ternario, alimentando la vía alostérica. Por el contrario, el intermedio con AD1 unido bloquea a Mdm2, forzando al sistema a usar la ruta competitiva en la que el TAD debe desprenderse completamente antes de que Mdm2 pueda unirse. Así, las formas microscópicas y fugaces que adopta el TAD durante la unión acaban dictando qué vía de intercambio a gran escala domina.
Conmutación rápida para decisiones de vida o muerte
Al juntar todos los números, los autores muestran que la vía alostérica mediante el complejo ternario puede soportar la mayor parte del tráfico entre los estados unidos a Mdm2 y a Taz2, especialmente cuando Mdm2 es abundante, como ocurre en las células. Esta ruta permite que p53 cambie rápidamente de estar retenido por Mdm2 a ser activado por Taz2 cuando cambian las condiciones, limitando al mismo tiempo la re-asociación excesiva de Mdm2. Las modificaciones químicas en el TAD que debilitan la unión a Mdm2 pueden desplazar el equilibrio de nuevo hacia la vía competitiva, ofreciendo otra capa de control. En términos sencillos, el estudio revela que un segmento proteico flexible utiliza su capacidad de cambiar de forma para construir un pequeño y temporal apretón de manos tridireccional que acelera el intercambio de una compañera por otra. Este mecanismo puede ser una forma general en que las regiones desordenadas de las proteínas gestionan redes de señalización complejas y ayudan a las células a responder rápidamente al estrés.
Cita: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Palabras clave: proteínas intrínsecamente desordenadas, regulación de p53, redes de unión de proteínas, conmutación alostérica, FRET de una sola molécula