Clear Sky Science · tr
IDP etkileşim ağının çeşitli bağlanma yollarıyla dinamik kontrolü
Hücreler Nasıl Hızlı Kararlar Alır
Hücrelerimizin içinde proteinler sürekli ortaklarını seçiyor, saniyenin kesirleri içinde bağlantılar kurup koparıyor. Bu seçimler bir hücrenin büyüyüp büyümeyeceğini, hasarı onarıp onaramayacağını ya da intihara benzer bir sürece girip girmeyeceğini belirleyebilir. Bu çalışma, tümör baskılayıcı p53’ten bir özellikle önemli protein segmentinin, p53 aktivitesini kapatan bir ortak ile onu etkinleştirmeye yardımcı olan bir diğer zıt ortak arasında nasıl hızla geçiş yaptığını inceliyor. Tek molekülleri gözleyerek, yazarlar bu geçişin tek bir basit rotayı izlemediğini, bunun yerine hücrelere strese karşı daha hızlı ve esnek yanıt verme olanağı sağlayan iki farklı yol kullandığını gösteriyor.
Şekil Değiştiren Bir Protein Merkezi
Hücrelerimizde birçok proteinin doğası gereği çözülebilir, düzensiz bölgeleri vardır; bunlara intrinsically disordered proteins (IDP) denir. Katı, iyi katlanmış proteinlerin aksine IDP’ler birçok farklı ortağa uyum sağlayabilir ve yoğun moleküler ağlarda merkez görevi görebilir. Ekip, p53’ün baş kısmındaki transaktivasyon domeni (TAD) üzerinde yoğunlaştı; klasik bir IDP segmenti olan bu bölge, AD1 ve AD2 adını taşıyan iki küçük alt bölge aracılığıyla birçok düzenleyici ile etkileşir. Ortaklardan biri olan Mdm2, p53’ün yıkımını teşvik eder ve hücre sağlıklı olduğunda düzeylerini düşük tutar. Diğer ortak Taz2 ise p53’ün stres yanıtı genlerini açmasına yardımcı olan büyük bir koaktivator kompleksinin parçasıdır. Taz2 hem AD1 hem AD2’ye bağlanabilirken Mdm2 yalnızca AD1’e bağlandığı için, p53 TAD tek bir esnek protein parçasının zıt etkileri olan ortakları nasıl idare edebileceğini sormak için ideal bir sistemdir.
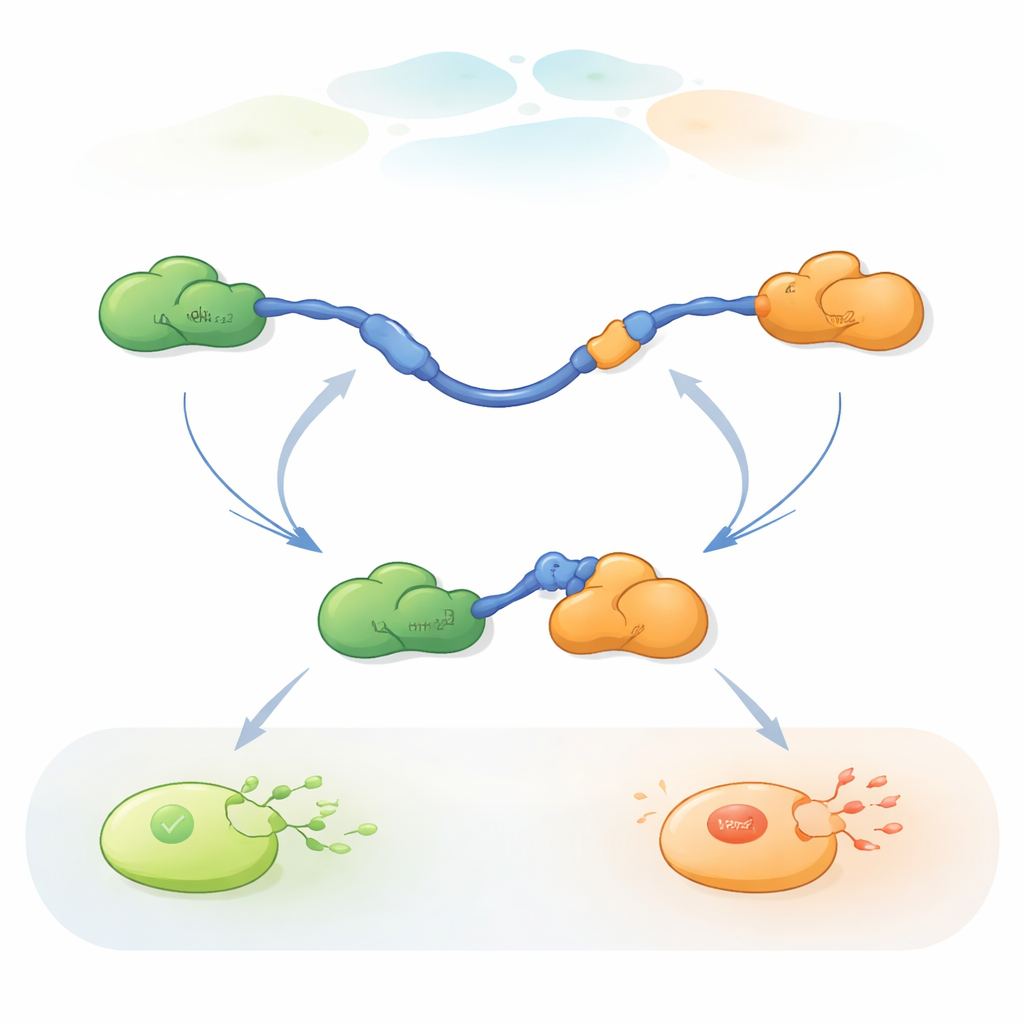
Tek Moleküllerin Ortak Değiştirişini İzlemek
Bu hızlı ortak değişimlerini izlemek için araştırmacılar, protein içi ve proteinler arası çok küçük mesafe değişikliklerini ölçen üç renkli tek-molekül FRET yöntemini kullandılar. Farklı renkli boyaları p53 TAD’nin uçlarına ve Taz2’ye bağlarken Mdm2’yi etiketlemediler. TAD’yi cam bir yüzeye immobilize edip ortakları kontrollü miktarlarda ekleyerek, bireysel p53 moleküllerinin zaman içinde Taz2 ve Mdm2’ye nasıl bağlandığını, ayrıldığını ve yeniden bağlandığını izleyebildiler. Floresans desenindeki değişimler, TAD’nin bağlı olmadığını, yalnızca Taz2’ye bağlı olduğunu, yalnızca Mdm2’ye bağlı olduğunu ya da kısa süreliğine her ikisine birden bağlanarak üç proteinli (ternary) bir kompleksi oluşturduğunu ortaya koydu.
Ortak Takasını Gerçekleştirmenin İki Yolu
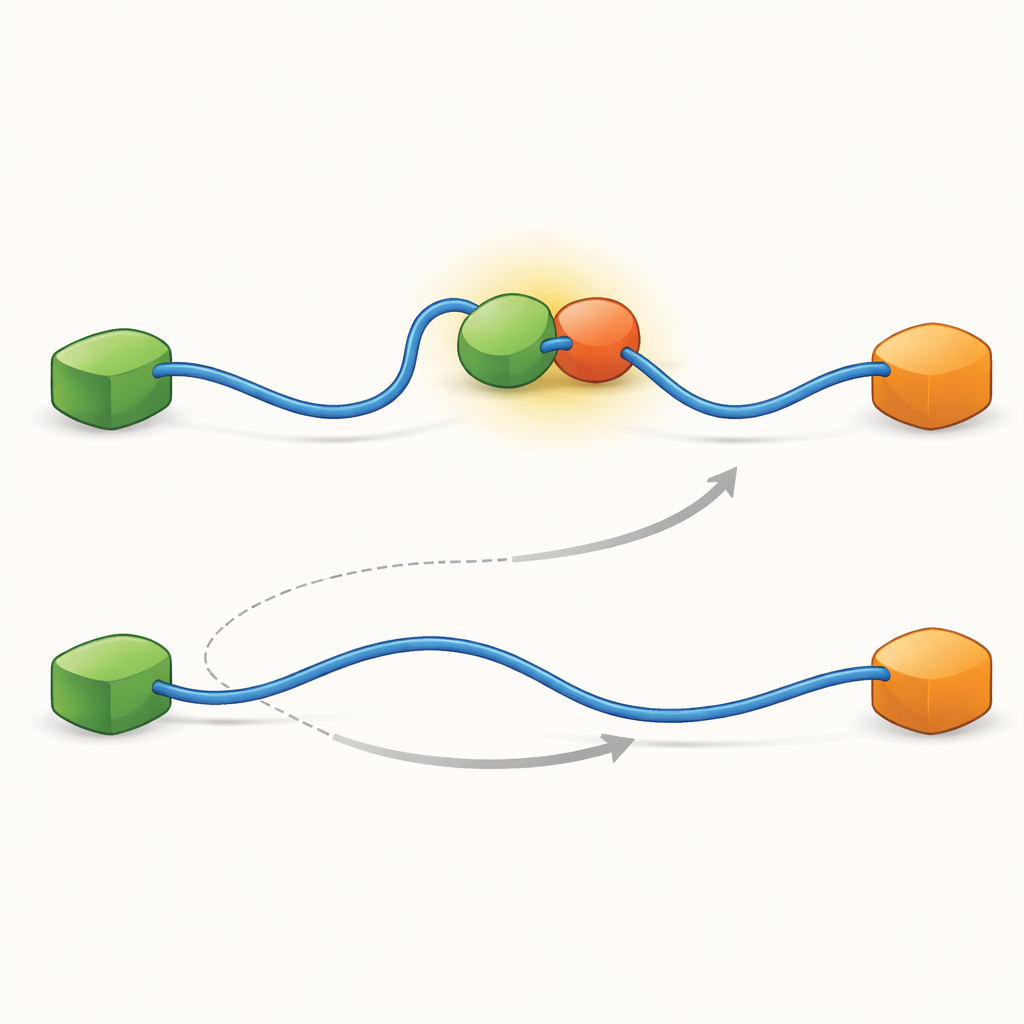
Kayıtlar, TAD’nin ortak değişimini niteliksel olarak iki farklı şekilde gerçekleştirebildiğini gösterdi. Rekabetçi yolda, TAD önce bir ortağından tamamen kopar, serbest ve esnek hale gelir ve ancak o zaman diğer ortağa bağlanır. Allosterik yolda ise TAD tam olarak ayrılmaz. Bunun yerine Taz2 ve Mdm2’nin aynı anda bağlandığı kısa ömürlü bir ternary kompleks oluşur; her biri TAD’nin farklı bir bölümünü tutar. Bu ortak durumdan bir ortak ayrılır ve diğeri kalır. TAD’nin her durumda ne kadar süre kaldığını ve geçişlerin ne sıklıkta gerçekleştiğini sayarak, yazarlar her adım için hızlılık sabitlerini çıkarabildiler ve farklı koşullar altında hangi yolun ne sıklıkta kullanıldığını nicelendirdiler.
Gizli Ara Adımlar Ağın Biçimini Belirler
Bu iki yolun nasıl mümkün olduğunu ortaya çıkarmak için ekip, tekniklerini çok yüksek zaman çözünürlüğüne kadar zorladı ve foton geliş olaylarını tek tek analiz etti. TAD’nin Taz2’ye bağlanma eylemine odaklandılar ve bu sürecin kendisinin bağlı olmayan ile tam bağlı durumlar arasında iki ayrı tür “geçiş yolu” izlediğini buldular — bağlanma sürecinde kısa süreli ara aşamalar. Bir türde Taz2 AD2’ye güçlü şekilde tutunurken AD1 serbest kalır; diğer türde ise Taz2 AD1’i daha güçlü şekilde tutar ve AD2 daha gevşektir. AD2’ye bağlı ara durum doğal olarak Mdm2’nin AD1’e sızmasına ve ternary kompleksin oluşmasına izin verir; bu da allosterik yolu besler. Buna karşılık AD1’e bağlı ara durum Mdm2’yi engeller ve sistemin TAD tamamen ayrılana kadar Mdm2’nin bağlanmasını engelleyerek rekabetçi yolu kullanmaya zorlar. Böylece TAD’nin bağlanma sırasında benimsediği mikroskobik, geçici şekiller hangi büyük ölçekli değişim yolunun baskın olacağını belirler.
Hayat veya Ölüm Kararları için Hızlı Anahtarlama
Tüm sayıları bir araya koyduklarında, yazarlar ternary kompleksli allosterik yolun, özellikle Mdm2 bol olduğunda (hücrelerde olduğu gibi), Mdm2-bağlı ve Taz2-bağlı durumlar arasındaki trafiğin çoğunu taşıyabileceğini gösteriyor. Bu yol, p53’ün Mdm2 tarafından tutulur haldeyken koşullar değiştiğinde Taz2 tarafından hızla etkinleştirilebilmesini sağlar ve aynı zamanda Mdm2’nin gereksiz yeniden bağlanmasını sınırlamaya yardımcı olur. TAD’ye kimyasal modifikasyonlar yapılarak Mdm2 bağlanmasının zayıflatılması, dengeyi rekabetçi yöne kaydırabilir ve başka bir kontrol katmanı sunar. Basitçe ifade etmek gerekirse, çalışma esnek bir protein segmentinin şekil değiştirme doğasını kullanarak, bir ortağın diğerine hızlıca değişmesini hızlandıran küçük, geçici üçlü bir tokalaşma oluşturduğunu ortaya koyuyor. Bu mekanizma, düzensiz protein bölgelerinin karmaşık sinyal ağlarını yönetme ve hücrelerin strese hızlı yanıt vermesine yardımcı olma biçimi olarak genel bir yol olabilir.
Atıf: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Anahtar kelimeler: doğası gereği düzensiz proteinler, p53 düzenlemesi, protein bağlanma ağları, allosterik anahtarlama, tek-molekül FRET