Clear Sky Science · he
בקרה דינמית על רשת אינטראקציות של חלבונים בלתי-מקובעים באמצעות מסלולי קשירה מגוונים
איך תאים מקבלים החלטות מהירות
בתוך התאים שלנו, חלבונים בוחרים באופן רציף שותפים, יוצרים ושוברים קשרים בשניות חלקיות. הבחירות האלה יכולות להכריע האם תא יגדל, יתקן נזק או ימות. המחקר בוחן כיצד קטע חלבון חשוב מהדיכא סרטן p53 עובר במהירות החלפה בין שני שותפים מנוגדים: אחד שמכבה את פעילות p53 ואחר שעוזר להפעילה. בעזרת צפייה במולקולות בודדות בפעולה הראו החוקרים שההחלפה הזו אינה נסמכת על מסלול פשוט יחיד, אלא עושה שימוש בשני מסלולים מובחנים שנותנים לתא דרך מהירה וגמישה יותר להגיב ללחץ.
ציר חלבוני שמשנה צורה
הרבה חלבונים בתאים שלנו כוללים אזורים רפויים ובלתי מקובעים הידועים כחלבונים באופן פנימית בלתי-מקובעים (IDPs). לא בדומה לחלבונים נוקשים ומקופלים היטב, IDP-ים יכולים לעצב את עצמם כדי להתאים לשותפים רבים, ופועלים כמרכזים ברשתות מולקולריות צפופות. הצוות התמקד בתחום האקטיבציה (TAD) בתחילת p53, קטע IDP קלאסי שמשתמש בשני אזורים קטנים, המכונים AD1 ו-AD2, כדי לקשור רגולטורים רבים. שותף אחד, Mdm2, מקדם פירוק של p53 ומחזיק את רמתו נמוכה כשהתא בריא. שותף אחר, Taz2, הוא חלק מקומפלקס קואקטיבטור גדול שעוזר ל-p53 להפעיל גנים בתגובה ללחץ. מאחר ש-Taz2 יכול לקשור גם את AD1 וגם את AD2 בעוד ש-Mdm2 קושר רק את AD1, ה-TAD של p53 מהווה מערכת אידיאלית לבדוק כיצד מקטע גמיש יחיד יכול לאזן בין שותפים שמניבים השפעות מנוגדות.
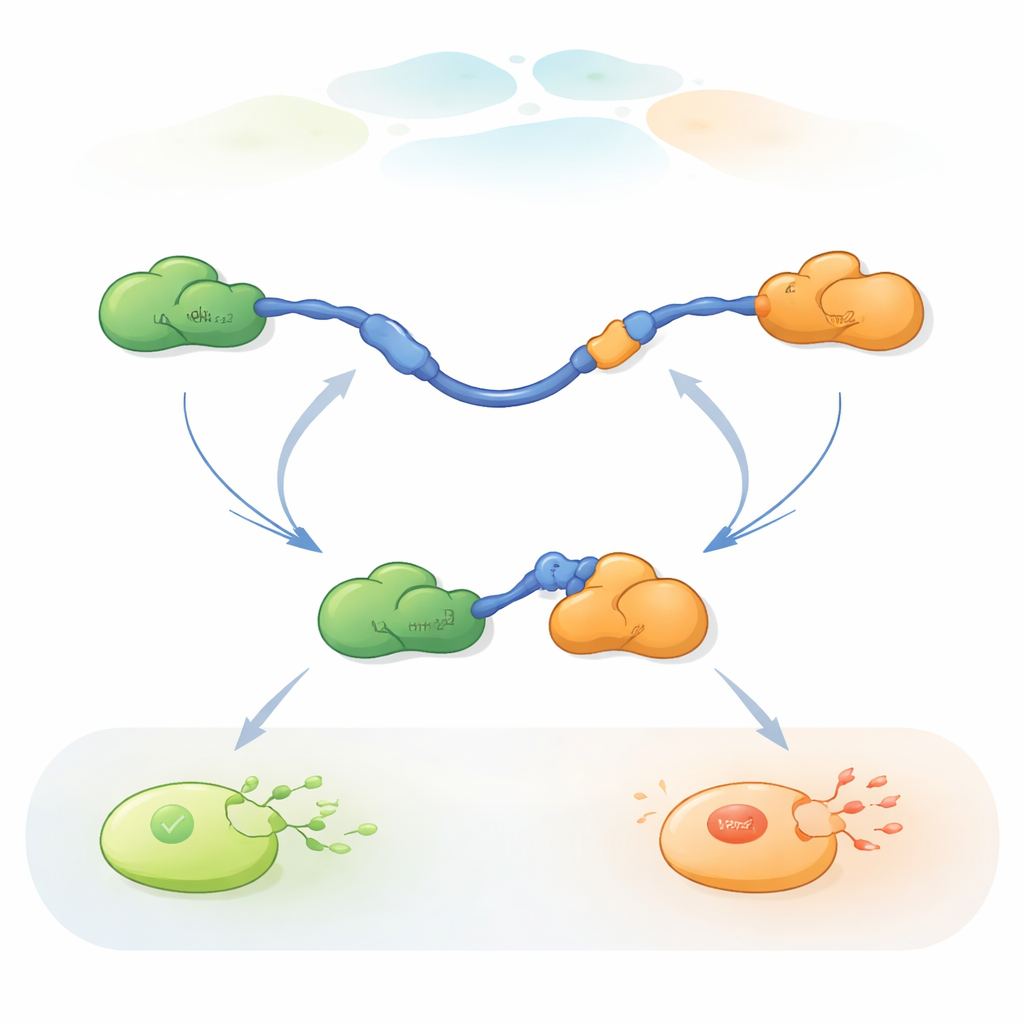
צפייה בהחלפות שותפים של מולקולות בודדות
כדי לעקוב אחרי החלפות השותפים המהירות הללו השתמשו החוקרים ב-FRET תלת-צבעוני של מולקולה בודדת, שיטה פלואורסצנטית המודדת שינויים זעירים במרחק בתוך ובין חלבונים. הם חיברו צבעים שונים לקצוות ה-TAD של p53 ול-Taz2, בעוד ש-Mdm2 נותר ללא תג. בהצמדת ה-TAD על משטח זכוכית ובהוספת השותפים בכמויות מבוקרות יכלו לצפות במולקולות p53 בודדות כשהן נקשרות, משתחררות ומשתלבות מחדש עם Taz2 ו-Mdm2 לאורך זמן. שינויים בדפוס הפלואורסצנציה חשפו האם ה-TAD לא קשור, קשור רק ל-Taz2, קשור רק ל-Mdm2, או קשור זמנית לשניהם בו זמנית במורכב שלושה-חלבונים (טרנרי).
שתי דרכים להחליף שותפים
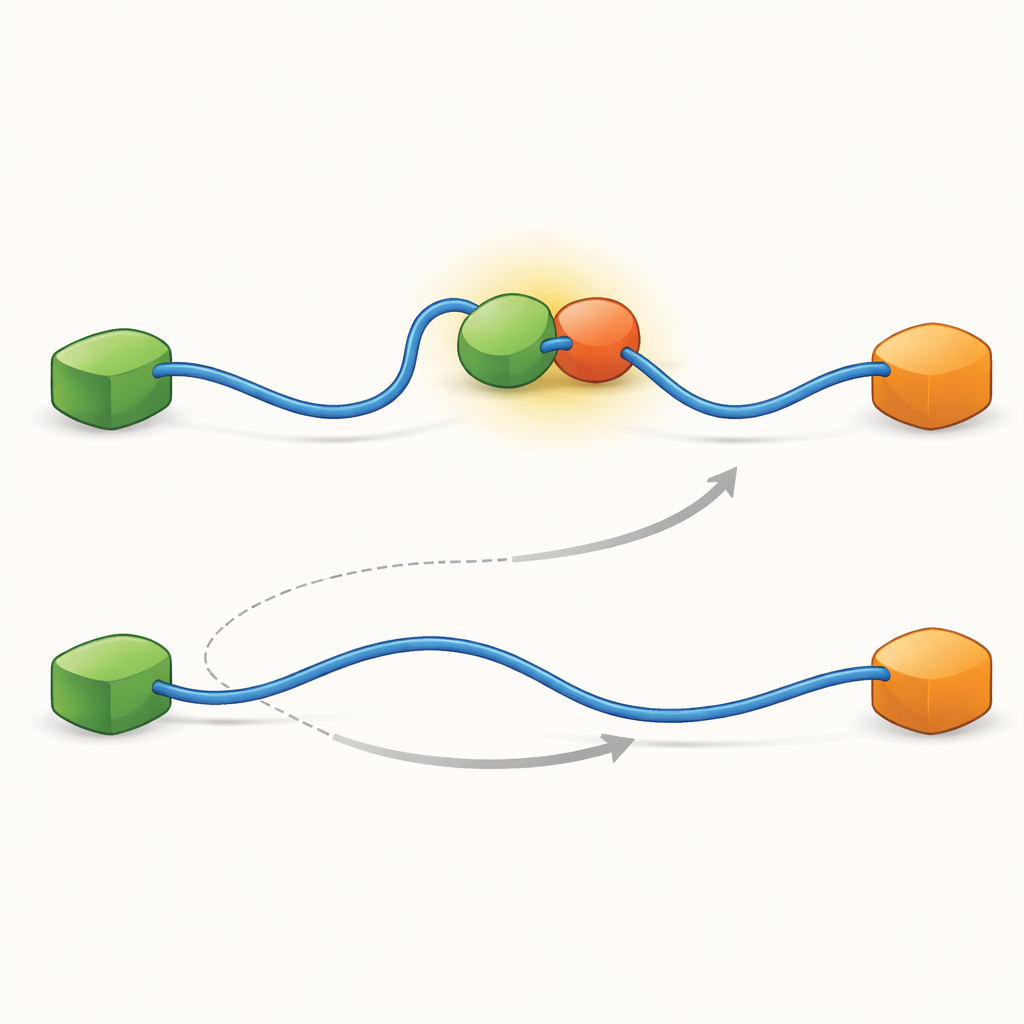
הרישומים הראו כי ה-TAD יכול להחליף שותפים בשתי דרכים איכותית שונות. בדרך התחרותית ה-TAD משחרר תחילה שותף באופן מלא, נעשה חופשי וגמיש, ורק אז נקשר לשותף השני. בדרך האלוסטרית ה-TAD אינו מתנתק לגמרי. במקום זאת הוא יוצר מורכב טרנרי קצר-חיים שבו Taz2 ו-Mdm2 קשורים בו זמנית, כל אחד תופס חלק שונה ב-TAD. ממצב משותף זה, אחד השותפים עוזב והשני נשאר. על ידי ספירת משכי הזמן שבהם ה-TAD שהה בכל מצב וכמה פעמים בוצעו המעברים, הצליחו המחברים לחלץ קבועי קצב לכל שלב ולכמת כמה לעתים כל מסלול מופעל בתנאים שונים.
שלבים בינוניים נסתרים מעצבים את הרשת
כדי לגלות מה מאפשר את שני המסלולים הללו דחפו הצוות את הטכניקה שלהם לרזולוציית זמן גבוהה מאוד וניתחו אירועי הגעה של פוטונים אחד-אחד. הם התמקדו במעשה קשירת ה-TAD ל-Taz2 ומצאו שתהליך זה עצמו עובר דרך שני סוגים מובחנים של «נתיבי מעבר»—שלבים בינוניים קצרים בין המצבים הלא קשור והקשור במלואם. בסוג אחד Taz2 אוחז בחוזקה ב-AD2 בעוד AD1 נשאר חופשי; בסוג האחר Taz2 מעורב יותר עם AD1 בעוד AD2 רפוי יותר. הביניים הקשור ל-AD2 מאפשר באופן טבעי ל-Mdm2 להחליק על AD1 וליצור את המורכב הטרנרי, מה שמזין את המסלול האלוסטרי. לעומת זאת, הביניים הקשור ל-AD1 חוסם את Mdm2 וכופה שימוש בדרך התחרותית שבה ה-TAD חייב להתנתק לחלוטין לפני ש-Mdm2 יוכל להקשר. לפיכך, הצורות המיקרוסקופיות והקצרות שה-TAD מאמץ במהלך הקשירה קובעות בסופו של דבר איזה מסלול החלפה בקנה מידה גדול ישלוט.
החלפה מהירה עבור בחירות של חיים או מוות
בחיבור כל המספרים יחד מראים המחברים כי המורכב הטרנרי והמסלול האלוסטרי יכולים לשאת רוב התנועה בין המצב הקשור ל-Mdm2 לבין המצב הקשור ל-Taz2, במיוחד כאשר Mdm2 בשפע כפי שקורה בתאים. נתיב זה מאפשר ל-p53 לעבור במהירות ממצב שבו הוא מעוכב על ידי Mdm2 למצב שבו הוא מופעל על ידי Taz2 כאשר התנאים משתנים, תוך הגבלת קשירה חוזרת מיותרת של Mdm2. שינויים כימיים ב-TAD שמחלישים את קשירת Mdm2 יכולים להזיז את האיזון חזרה אל המסלול התחרותי ולהעניק שכבת בקרה נוספת. בפשטות, המחקר מגלה שמקטע חלבון גמיש משתמש באופיו המשנה-צורה כדי לבנות לחימה של שלוש דרכים זמנית שמאיצה את החלפת השותפים. מנגנון זה עשוי להיות דרך כללית שבה אזורים בלתי-מקובעים של חלבון מנהלים רשתות איתות מורכבות ועוזרים לתאים להגיב במהירות ללחץ.
ציטוט: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
מילות מפתח: חלבונים באופן פנימי בלתי-מקובעים, ויסות p53, רשתות קשירת חלבונים, הסבה אלוסטרית, FRET מולקולארי יחיד