Clear Sky Science · nl
Dynamische controle van het IDP-interactienetwerk via diverse bindingsroutes
Hoe cellen snelle beslissingen nemen
Binnen onze cellen kiezen eiwitten constant partners, waarbij verbindingen binnen fracties van een seconde worden gevormd en verbroken. Deze keuzes kunnen bepalen of een cel groeit, schade herstelt of zichzelf vernietigt. Deze studie onderzoekt hoe een bijzonder belangrijk eiwitsequentiefragment van de tumorsuppressor p53 snel schakelt tussen twee tegengestelde partners: de ene die p53-activiteit uitschakelt en de andere die helpt deze te activeren. Door individuele moleculen in actie te volgen laten de auteurs zien dat dit schakelen niet via één eenvoudige route verloopt, maar twee onderscheiden paden gebruikt die de cel een snellere en flexibelere reactie op stress geven.
Een van vorm veranderend eiwithub
Veel eiwitten in onze cellen hebben slappe, ongestructureerde regio’s die bekendstaan als intrinsiek ongeordende eiwitten (IDP’s). In tegenstelling tot stijve, goedgevouwen eiwitten kunnen IDP’s zich vormen naar verschillende partners en fungeren ze als knooppunten in drukke moleculaire netwerken. Het team richtte zich op het transactivatiedomein (TAD) aan het begin van p53, een klassiek IDP-segment dat twee kleine regio’s gebruikt, genoemd AD1 en AD2, om met vele regulatoren te interageren. Eén partner, Mdm2, bevordert de afbraak van p53 en houdt de niveaus laag wanneer de cel gezond is. Een andere partner, Taz2, maakt deel uit van een groot coactivatorcomplex dat p53 helpt stress-responsgenen aan te zetten. Omdat Taz2 zowel aan AD1 als AD2 kan binden terwijl Mdm2 alleen AD1 bindt, is p53 TAD een ideaal systeem om te onderzoeken hoe één flexibel eiwitdeel met tegengestelde partners kan jongleren.
Individuele moleculen zien van partner wisselen
Om deze snelle partnerwissels te volgen gebruikten de onderzoekers driekleurige single-molecule FRET, een fluorescentiemethode die zeer kleine afstandsveranderingen binnen en tussen eiwitten meet. Ze bevestigden verschillende gekleurde kleurstoffen aan de uiteinden van p53 TAD en aan Taz2, terwijl Mdm2 niet werd gelabeld. Door TAD op een glasoppervlak te immobiliseren en de partners in gecontroleerde hoeveelheden toe te voegen, konden ze individuele p53-moleculen observeren terwijl ze Taz2 en Mdm2 binden, loslaten en opnieuw binden in de tijd. Veranderingen in het fluorescentiepatroon toonden of TAD ongebonden was, alleen aan Taz2 gebonden, alleen aan Mdm2 gebonden, of kort aan beide tegelijk in een drie-eiwit (ternair) complex.
Twee manieren om partners te ruilen
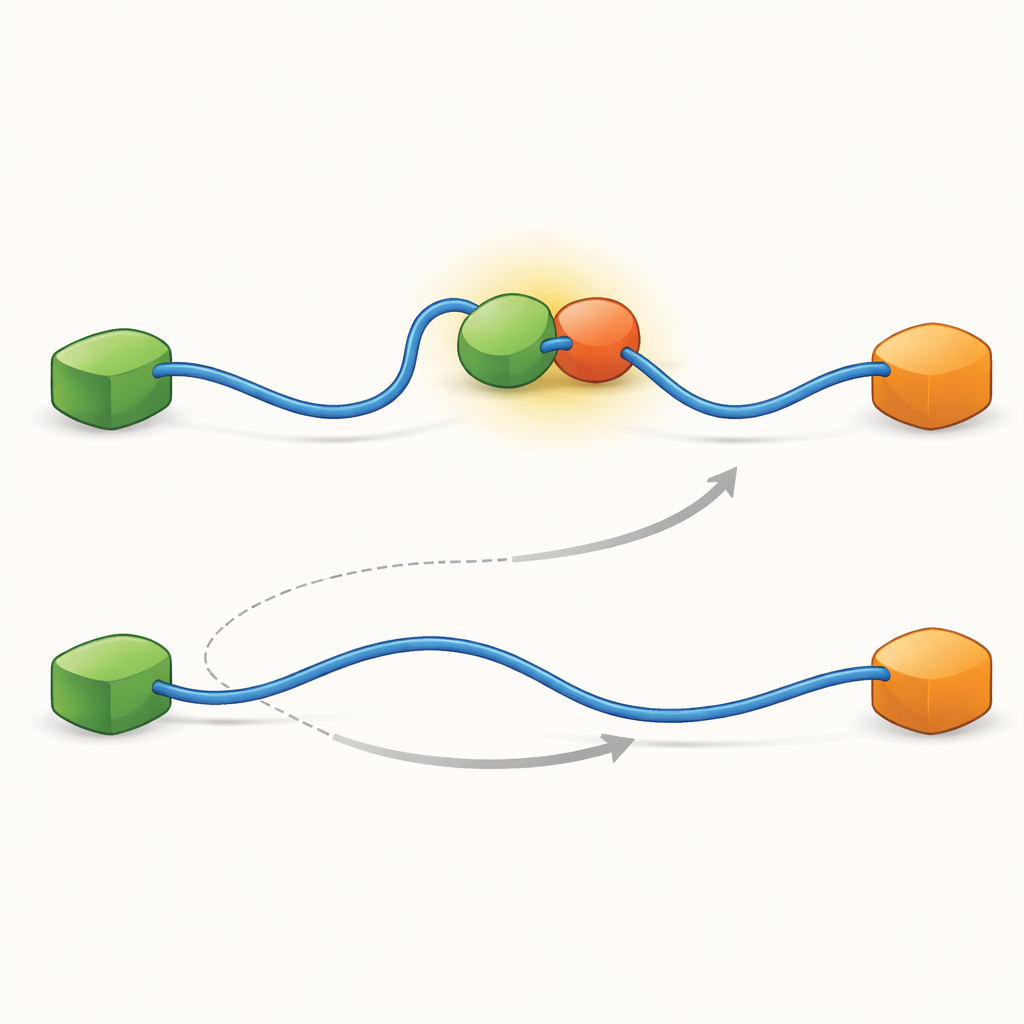
De opnames toonden aan dat TAD partners op twee kwalitatief verschillende manieren kan uitwisselen. In het competitieve pad laat TAD eerst volledig één partner los, wordt vrij en flexibel, en bindt pas daarna de andere partner. In het allosterische pad detacheert TAD niet volledig. In plaats daarvan vormt het een kortlevend ternair complex waarin Taz2 en Mdm2 gelijktijdig gebonden zijn, elk zich vastgrijpend aan een ander deel van TAD. Vanuit deze gezamenlijke toestand vertrekt de ene partner en blijft de andere over. Door te tellen hoe lang TAD in elke toestand verbleef en hoe vaak overgangen plaatsvonden, konden de auteurs snelheidconstanten voor elke stap afleiden en kwantificeren hoe vaak elk pad wordt gekozen onder verschillende omstandigheden.
Verborgen tussenstappen vormen het netwerk
Om te achterhalen wat deze twee paden mogelijk maakt, bracht het team hun techniek naar zeer hoge tijdresolutie en analyseerden ze foton-aankomstgebeurtenissen één voor één. Ze concentreerden zich op de daad van TAD-bindig aan Taz2 en vonden dat dit proces zelf twee verschillende soorten “overgangspaden” volgt—korte tussenfasen tussen ongebonden en volledig gebonden toestanden. In het ene geval houdt Taz2 sterk vast aan AD2 terwijl AD1 vrij blijft; in het andere engageert Taz2 sterker met AD1 terwijl AD2 losser is. Het AD2-gebonden intermediair laat het toe dat Mdm2 op AD1 inschuift en het ternaire complex vormt, wat het allosterische pad voedt. Het AD1-gebonden intermediair blokkeert daarentegen Mdm2, waardoor het systeem het competitieve traject moet volgen waarbij TAD eerst volledig loskomt voordat Mdm2 kan binden. Zo bepalen de microscopische, vluchtige vormen die TAD tijdens binden aanneemt welke grootschalige uitwisselingsweg domineert.
Snelle schakeling voor levens-of-doodkeuzes
Als je alle cijfers bij elkaar optelt, tonen de auteurs aan dat het ternaire, allosterische pad het grootste deel van het verkeer tussen Mdm2-gebonden en Taz2-gebonden toestanden kan dragen, vooral wanneer Mdm2 overvloedig aanwezig is, zoals in cellen het geval is. Deze route stelt p53 in staat snel te schakelen van onder controle gehouden door Mdm2 naar geactiveerd door Taz2 wanneer de omstandigheden veranderen, terwijl overmatige herkoppeling van Mdm2 toch wordt beperkt. Chemische modificaties aan TAD die de Mdm2-binding verzwakken kunnen het evenwicht terug verplaatsen naar het competitieve pad, wat een extra controlelaag biedt. Simpel gezegd onthult de studie dat een flexibel eiwitsegment zijn vormveranderende aard gebruikt om een kleine, tijdelijke driehanddruk te vormen die het ruilen van de ene partner voor de andere versnelt. Dit mechanisme kan een algemene manier zijn waarop ongeordende eiwitreïgioenen complexe signaalnetwerken beheren en cellen helpen snel op stress te reageren.
Bronvermelding: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Trefwoorden: intrinsiek ongeordende eiwitten, p53-regulatie, eiwitbindingsnetwerken, allosterische schakeling, single-molecule FRET