Clear Sky Science · fr
Contrôle dynamique du réseau d’interactions des protéines désordonnées via des voies de liaison variées
Comment les cellules prennent des décisions rapides
À l’intérieur de nos cellules, les protéines choisissent sans cesse des partenaires, formant et rompant des liaisons en une fraction de seconde. Ces choix peuvent déterminer si une cellule se développe, répare des dommages ou s’autodétruit. Cette étude examine comment un segment de protéine particulièrement important du suppresseur de tumeur p53 bascule rapidement entre deux partenaires opposés : l’un qui éteint l’activité de p53 et l’autre qui la stimule. En observant des molécules uniques en action, les auteurs montrent que ce basculement ne suit pas une unique voie simple, mais emprunte au contraire deux trajectoires distinctes qui offrent à la cellule une réponse plus rapide et plus flexible au stress.
Un centre protéique qui change de forme
Beaucoup de protéines dans nos cellules possèdent des régions souples et non structurées appelées protéines intrinsèquement désordonnées (IDP). À la différence des protéines rigides et bien repliées, les IDP peuvent se mouler pour convenir à de nombreux partenaires, jouant le rôle de nœuds dans des réseaux moléculaires denses. L’équipe s’est concentrée sur le domaine d’activation transcriptionnelle (TAD) situé au début de p53, un segment classique d’IDP qui utilise deux petites régions, appelées AD1 et AD2, pour interagir avec de nombreux régulateurs. Un partenaire, Mdm2, favorise la dégradation de p53 et maintient ses niveaux bas lorsque la cellule est en bonne santé. Un autre partenaire, Taz2, fait partie d’un large complexe coactivateur qui aide p53 à activer les gènes de réponse au stress. Parce que Taz2 peut se lier à la fois à AD1 et AD2 tandis que Mdm2 ne se lie qu’à AD1, le TAD de p53 constitue un système idéal pour étudier comment un fragment protéique flexible peut jongler avec des partenaires aux effets opposés.
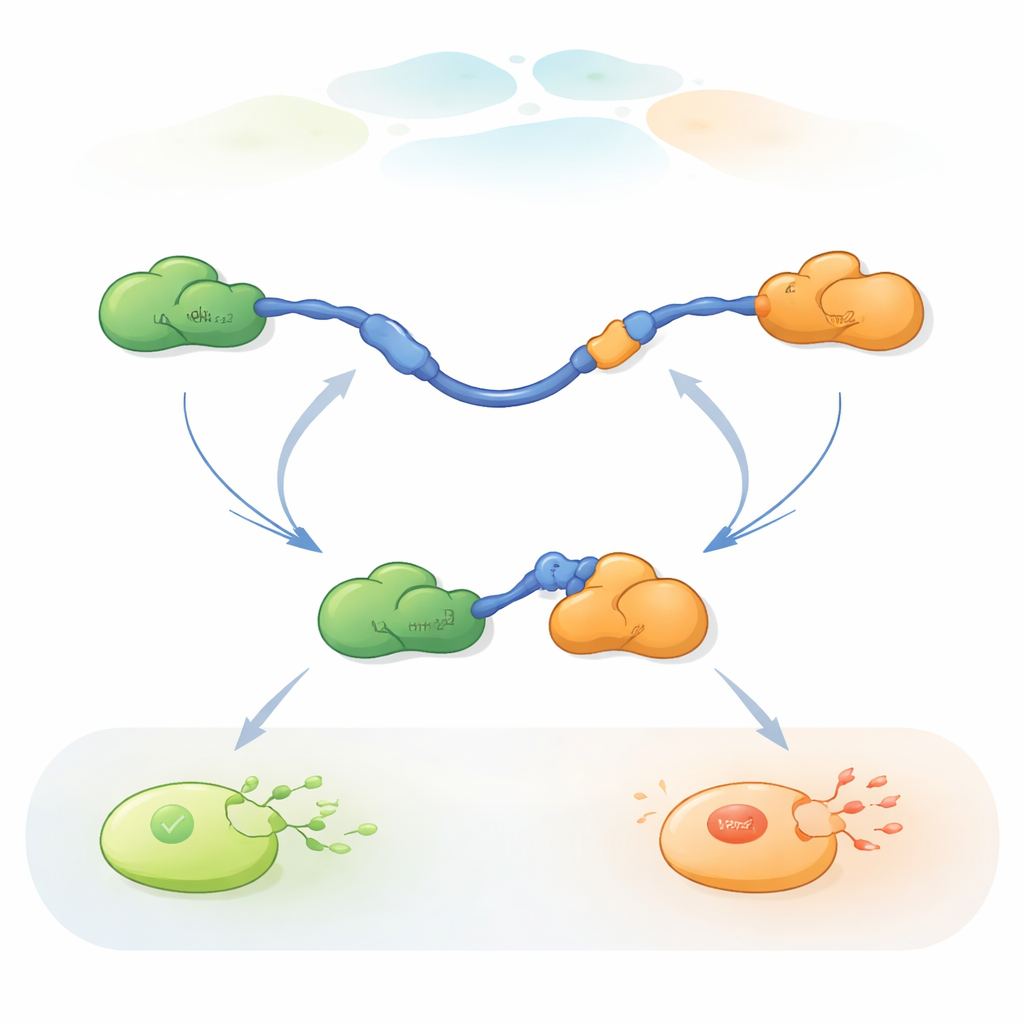
Observer des molécules uniques échanger de partenaires
Pour suivre ces échanges rapides de partenaires, les chercheurs ont utilisé le FRET tricolore monomoléculaire, une méthode de fluorescence qui mesure de très petites variations de distance à l’intérieur et entre protéines. Ils ont fixé des colorants de couleurs différentes aux extrémités du TAD de p53 et à Taz2, tout en laissant Mdm2 non marqué. En immobilisant le TAD sur une surface de verre et en ajoutant les partenaires en quantités contrôlées, ils ont pu observer des molécules de p53 individuelles se lier, se détacher et se relier à Taz2 et Mdm2 au fil du temps. Les changements dans le schéma de fluorescence ont révélé si le TAD était non lié, lié uniquement à Taz2, lié uniquement à Mdm2, ou brièvement lié aux deux simultanément dans un complexe ternaire à trois protéines.
Deux manières d’échanger un partenaire
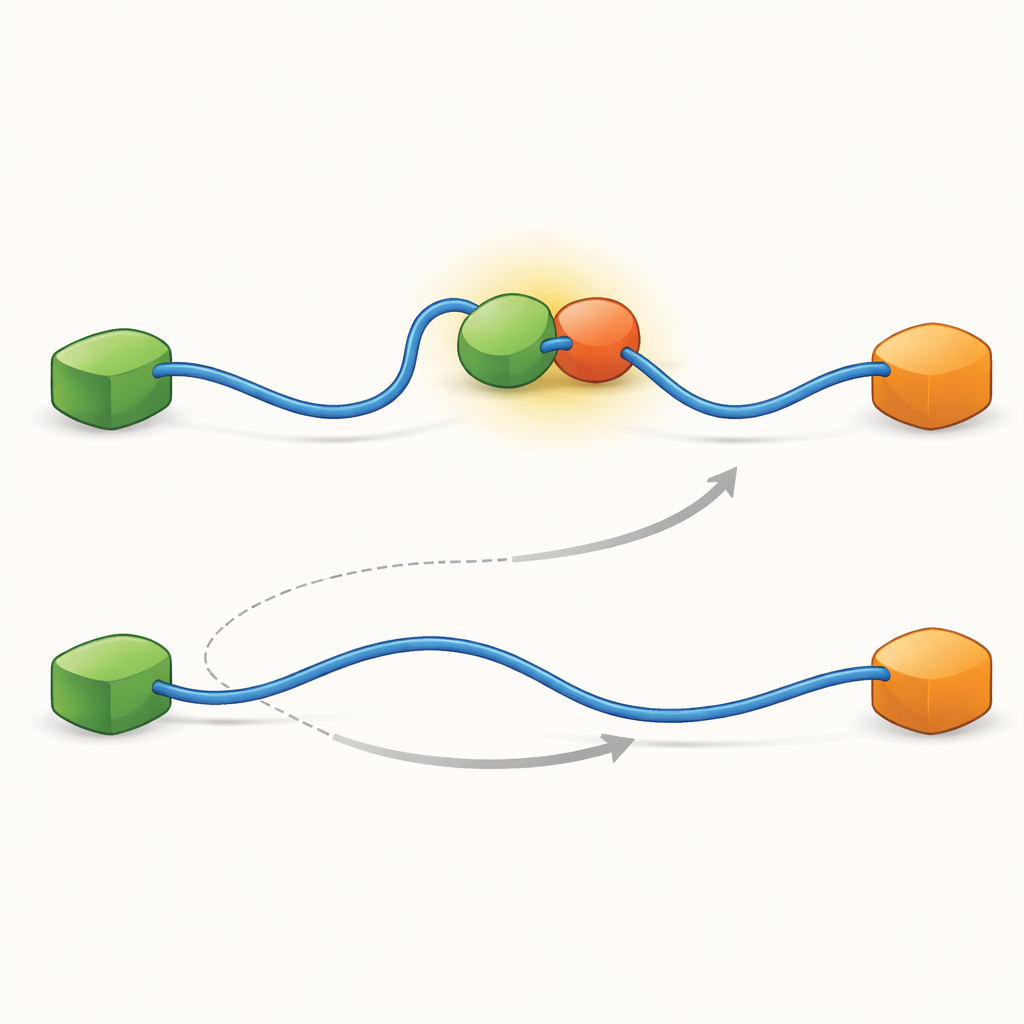
Les enregistrements ont montré que le TAD peut échanger ses partenaires de deux façons qualitativement différentes. Dans la voie compétitive, le TAD lâche d’abord complètement un partenaire, devenant libre et flexible, puis ne se lie qu’ensuite à l’autre partenaire. Dans la voie allostérique, le TAD ne se détache pas entièrement. Il forme plutôt un complexe ternaire de courte durée où Taz2 et Mdm2 sont tous deux liés en même temps, chacun s’attachant à une partie différente du TAD. À partir de cet état conjoint, un partenaire part et l’autre reste. En comptant combien de temps le TAD restait dans chaque état et la fréquence des transitions, les auteurs ont pu extraire des constantes de vitesse pour chaque étape et quantifier la fréquence d’utilisation de chaque voie selon les conditions.
Des étapes intermédiaires cachées façonnent le réseau
Pour comprendre ce qui rend ces deux voies possibles, l’équipe a poussé sa technique à très haute résolution temporelle et analysé les arrivées de photons une par une. Ils se sont concentrés sur l’acte d’association du TAD à Taz2 et ont constaté que ce processus suit lui-même deux types distincts de « chemins de transition » — des étapes intermédiaires brèves entre les états non lié et entièrement lié. Dans un type, Taz2 tient fortement à AD2 tandis qu’AD1 reste libre ; dans l’autre, Taz2 s’engage davantage sur AD1 tandis qu’AD2 est plus lâche. L’intermédiaire lié à AD2 permet naturellement à Mdm2 de se glisser sur AD1 et de créer le complexe ternaire, alimentant la voie allostérique. En revanche, l’intermédiaire lié à AD1 bloque Mdm2, forçant le système à emprunter la voie compétitive où le TAD doit se détacher complètement avant que Mdm2 puisse se lier. Ainsi, les formes microscopiques et fugaces que le TAD adopte pendant la liaison finissent par dicter quelle voie d’échange à grande échelle domine.
Un basculement rapide pour des choix vitaux
En réunissant toutes les mesures, les auteurs montrent que la voie allostérique passant par le complexe ternaire peut transporter la majeure partie du trafic entre les états liés à Mdm2 et à Taz2, en particulier lorsque Mdm2 est abondant, comme c’est le cas en cellule. Cette voie permet à p53 de passer rapidement d’un état d’inhibition par Mdm2 à un état d’activation par Taz2 lorsque les conditions changent, tout en limitant la recapture excessive par Mdm2. Des modifications chimiques du TAD qui affaiblissent la liaison à Mdm2 peuvent faire basculer l’équilibre vers la voie compétitive, offrant une couche de contrôle supplémentaire. En termes simples, l’étude révèle qu’un segment protéique flexible utilise sa nature changeante pour établir une petite poignée de main tertiaire temporaire qui accélère l’échange d’un partenaire contre un autre. Ce mécanisme pourrait être une façon générale pour les régions protéiques désordonnées de gérer des réseaux de signalisation complexes et d’aider les cellules à répondre rapidement au stress.
Citation: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Mots-clés: protéines intrinsèquement désordonnées, régulation de p53, réseaux de liaison des protéines, basculement allostérique, FRET monomoléculaire