Clear Sky Science · pt
Controle dinâmico da rede de interações de IDPs por vias de ligação diversas
Como as Células Tomam Decisões Rápidas
Dentro das nossas células, proteínas estão constantemente escolhendo parceiros, formando e rompendo conexões em frações de segundo. Essas escolhas podem determinar se uma célula cresce, repara danos ou se autodestroi. Este estudo investiga como um segmento proteico particularmente importante do supressor de tumor p53 alterna rapidamente entre dois parceiros opostos: um que desliga a atividade de p53 e outro que a ajuda a ativar. Ao observar moléculas individuais em ação, os autores mostram que essa troca não segue uma única rota simples, mas utiliza duas vias distintas que conferem às células uma maneira mais rápida e flexível de responder ao estresse.
Um Hub Proteico que Muda de Forma
Muitas proteínas em nossas células têm regiões frouxas e sem estrutura definida conhecidas como proteínas intrinsecamente desordenadas (IDPs). Ao contrário de proteínas rígidas e bem dobradas, as IDPs podem se moldar para acomodar muitos parceiros diferentes, atuando como hubs em redes moleculares congestionadas. A equipe se concentrou no domínio de transativação (TAD) no início de p53, um segmento clássico de IDP que usa duas pequenas regiões, chamadas AD1 e AD2, para interagir com vários reguladores. Um parceiro, Mdm2, promove a degradação de p53 e mantém seus níveis baixos quando a célula está saudável. Outro parceiro, Taz2, faz parte de um grande complexo coativador que ajuda p53 a ativar genes de resposta ao estresse. Como Taz2 pode se ligar tanto a AD1 quanto a AD2, enquanto Mdm2 liga-se apenas a AD1, o TAD de p53 é um sistema ideal para investigar como um segmento flexível de proteína pode conciliar parceiros com efeitos opostos.
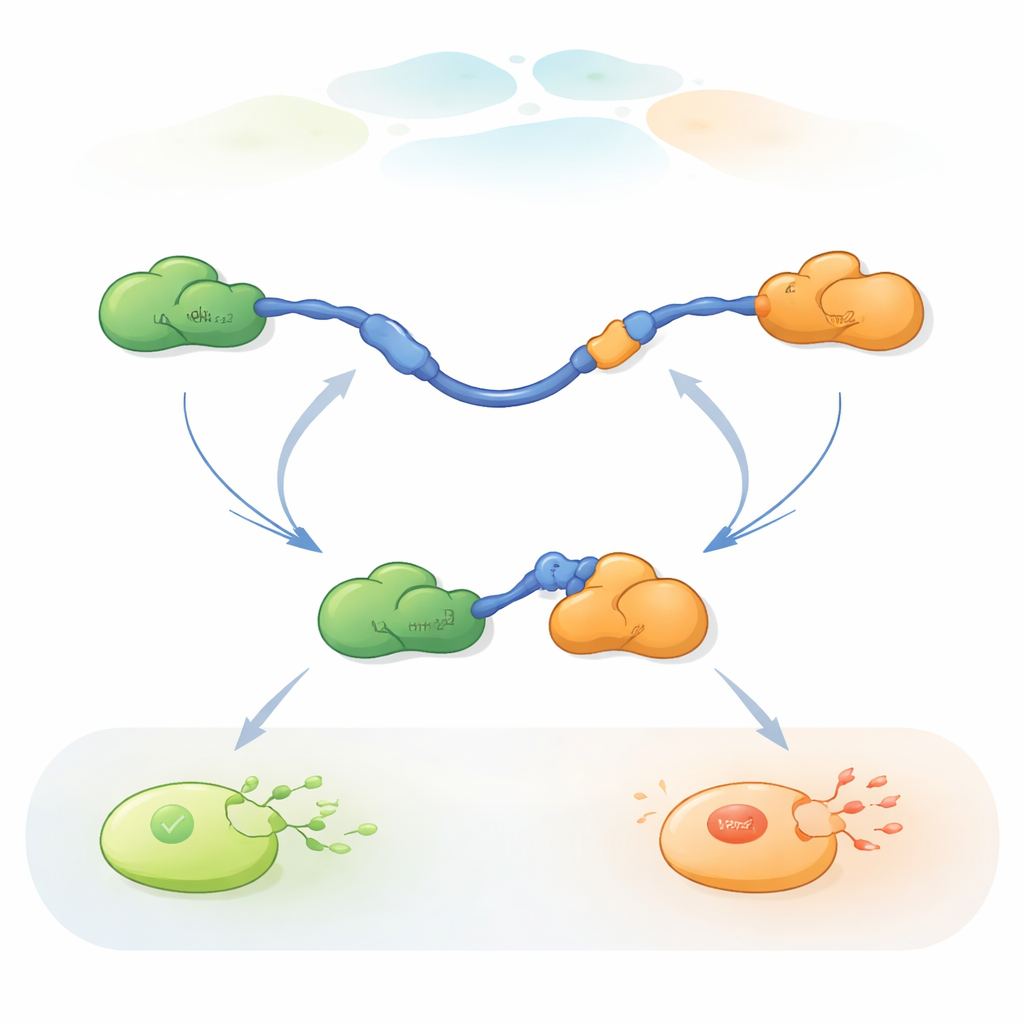
Observando Moléculas Individuais Trocarem de Parceiro
Para acompanhar essas trocas rápidas de parceiros, os pesquisadores usaram FRET de molécula única com três cores, um método de fluorescência que mede pequenas mudanças de distância dentro e entre proteínas. Eles acoplaram corantes de cores diferentes às extremidades do TAD de p53 e ao Taz2, enquanto deixaram o Mdm2 não marcado. Ao imobilizar o TAD em uma superfície de vidro e adicionar os parceiros em quantidades controladas, puderam observar moléculas individuais de p53 enquanto se ligavam, liberavam e se religavam a Taz2 e Mdm2 ao longo do tempo. Mudanças no padrão de fluorescência revelaram se o TAD estava não ligado, ligado apenas a Taz2, ligado apenas a Mdm2, ou brevemente ligado a ambos ao mesmo tempo em um complexo ternário de três proteínas.
Dois Jeitos de Trocar de Parceiro
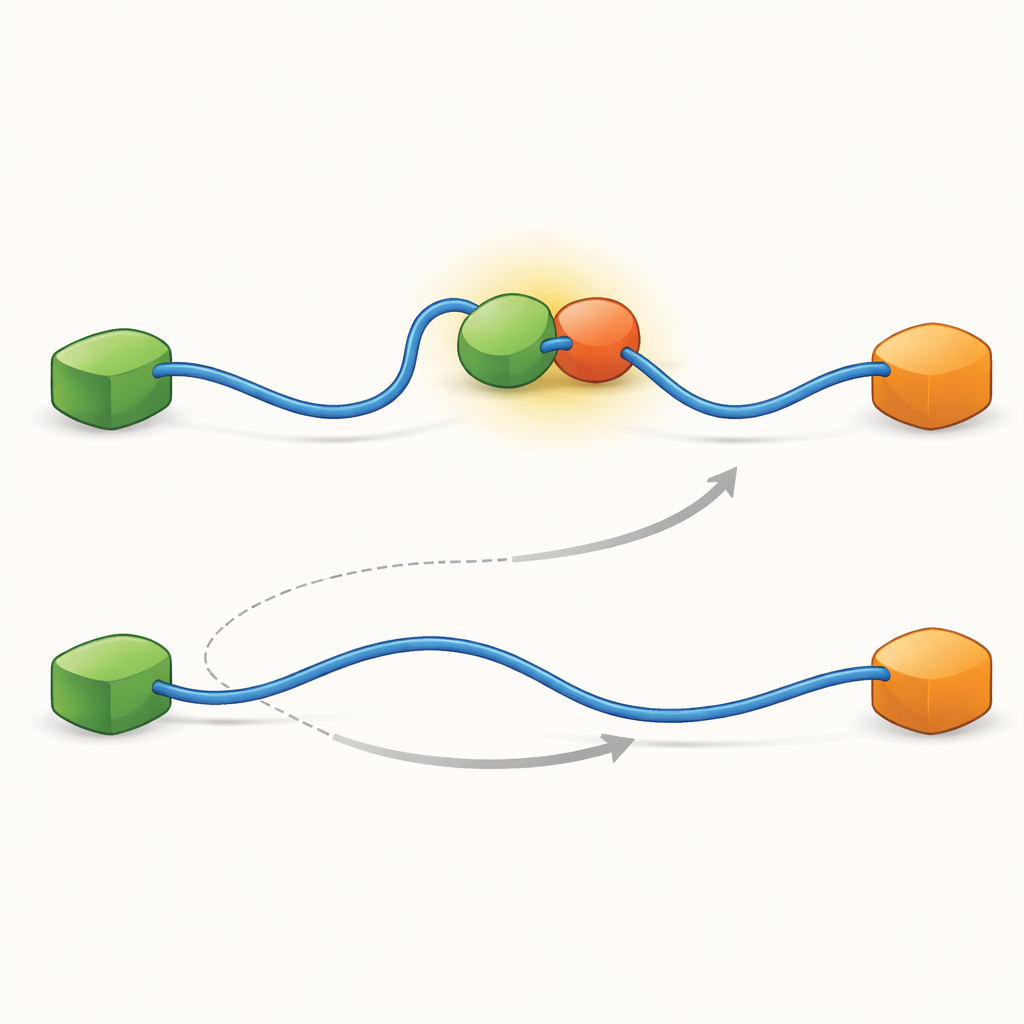
As gravações mostraram que o TAD pode trocar de parceiros de duas maneiras qualitativamente diferentes. Na via competitiva, o TAD primeiro se desprende completamente de um parceiro, tornando-se livre e flexível, e só então se liga ao outro parceiro. Na via alostérica, o TAD não se desliga totalmente. Em vez disso, forma um complexo ternário de curta duração em que Taz2 e Mdm2 estão ambos ligados ao mesmo tempo, cada um segurando uma parte diferente do TAD. A partir desse estado conjunto, um parceiro sai e o outro permanece. Contando quanto tempo o TAD permaneceu em cada estado e com que frequência ocorreram as transições, os autores puderam extrair constantes de taxa para cada etapa e quantificar com que frequência cada via é utilizada sob diferentes condições.
Etapas Intermediárias Ocultas Moldam a Rede
Para descobrir o que torna essas duas vias possíveis, a equipe levou sua técnica a resolução temporal muito alta e analisou os eventos de chegada de fótons um a um. Eles se concentraram no ato de ligação do TAD ao Taz2 e descobriram que esse processo em si segue dois tipos distintos de “caminhos de transição” — estágios intermediários breves entre os estados não ligado e totalmente ligado. Em um tipo, Taz2 prende-se fortemente a AD2 enquanto AD1 permanece livre; no outro, Taz2 envolve AD1 mais fortemente enquanto AD2 fica mais solto. O intermediário ligado a AD2 permite naturalmente que Mdm2 se encaixe em AD1 e crie o complexo ternário, alimentando a via alostérica. Por contraste, o intermediário ligado a AD1 bloqueia o Mdm2, forçando o sistema a usar a rota competitiva em que o TAD deve se desprender completamente antes que Mdm2 possa ligar-se. Assim, as formas microscópicas e fugazes que o TAD adota durante a ligação acabam ditando qual via de troca em grande escala predomina.
Trocas Rápidas para Decisões de Vida ou Morte
Consolidando todos os números, os autores mostram que o complexo ternário e a via alostérica podem conduzir a maior parte do tráfego entre os estados ligados a Mdm2 e Taz2, especialmente quando Mdm2 é abundante, como é nas células. Essa rota permite que p53 mude rapidamente de estar reprimido por Mdm2 para ser ativado por Taz2 quando as condições mudam, ao mesmo tempo em que limita o reatamento excessivo de Mdm2. Modificações químicas no TAD que enfraquecem a ligação ao Mdm2 podem deslocar o equilíbrio de volta para a via competitiva, oferecendo outra camada de controle. Em termos simples, o estudo revela que um segmento proteico flexível usa sua natureza mutável para formar um pequeno e temporário aperto triplo que acelera a troca de um parceiro por outro. Esse mecanismo pode ser uma forma generalizada pela qual regiões desordenadas de proteínas gerenciam redes de sinalização complexas e ajudam as células a responder rapidamente ao estresse.
Citação: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Palavras-chave: proteínas intrinsecamente desordenadas, regulação de p53, redes de ligação de proteínas, comutação alostérica, FRET de molécula única