Clear Sky Science · de
Dynamische Kontrolle des IDP-Interaktionsnetzwerks über verschiedene Bindungswege
Wie Zellen schnelle Entscheidungen treffen
In unseren Zellen wählen Proteine ständig Partner aus und gehen Verbindungen in Bruchteilen einer Sekunde ein oder lösen sie wieder. Diese Entscheidungen können bestimmen, ob eine Zelle wächst, Schäden repariert oder sich selbst zerstört. Diese Studie untersucht, wie ein besonders wichtiger Abschnitt des Tumorsuppressors p53 rasch zwischen zwei entgegengesetzten Partnern wechselt: einem, der die p53-Aktivität abschaltet, und einem anderem, der sie aktiviert. Durch Beobachtung einzelner Moleküle in Aktion zeigen die Autoren, dass dieser Wechsel keinem einzigen einfachen Weg folgt, sondern stattdessen zwei unterschiedliche Pfade nutzt, die Zellen eine schnellere und flexiblere Reaktion auf Stress ermöglichen.
Ein formwandelnder Proteinknoten
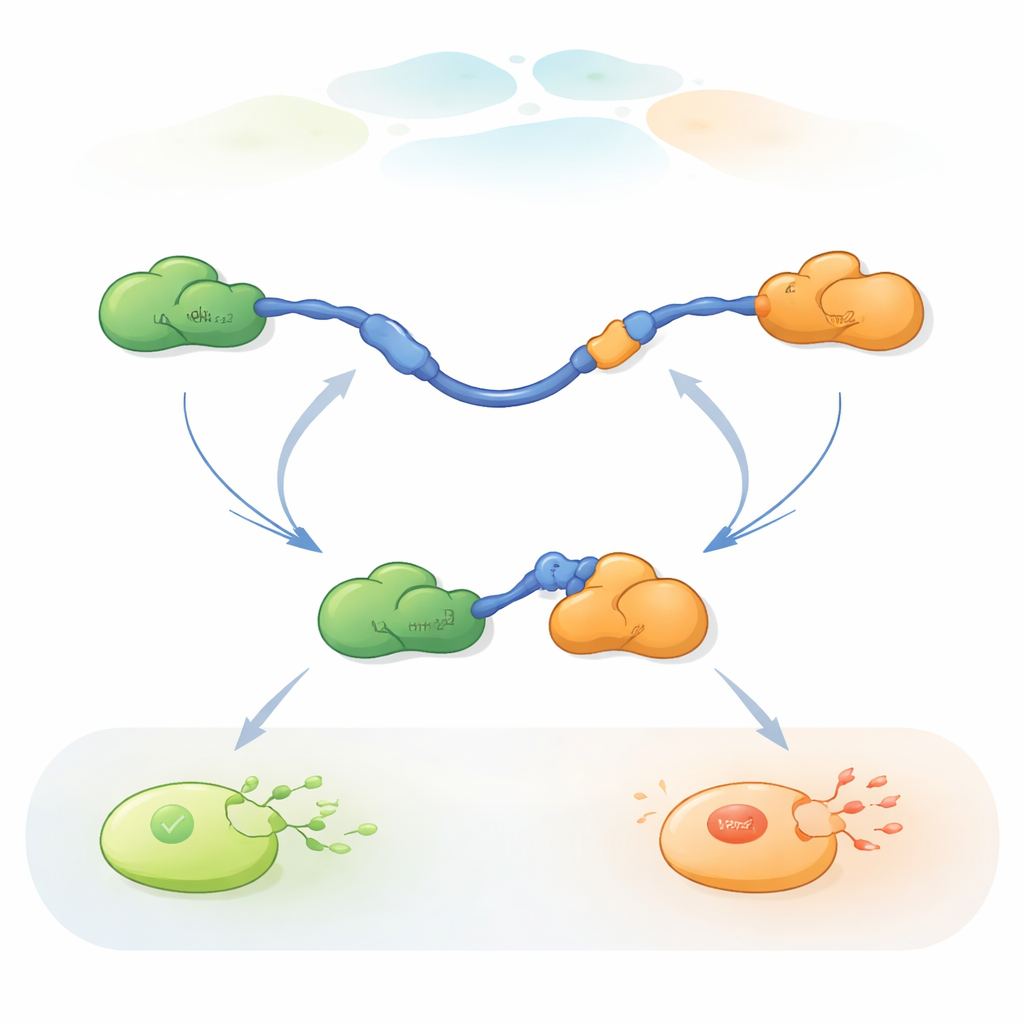
Viele Proteine in unseren Zellen besitzen flexible, ungeordnete Regionen, die als intrinsisch ungeordnete Proteine (IDPs) bezeichnet werden. Im Gegensatz zu starren, gut gefalteten Proteinen können sich IDPs an viele verschiedene Partner anpassen und fungieren so als Knotenpunkte in dicht gepackten molekularen Netzwerken. Das Team konzentrierte sich auf die Transaktivierungsdomäne (TAD) am Beginn von p53, ein klassisches IDP-Segment, das zwei kleine Regionen nutzt, genannt AD1 und AD2, um mit zahlreichen Regulatoren zu interagieren. Ein Partner, Mdm2, fördert den Abbau von p53 und hält dessen Spiegel gering, wenn die Zelle gesund ist. Ein anderer Partner, Taz2, gehört zu einem großen Coaktivator-Komplex, der p53 dabei hilft, Stressantwort-Gene einzuschalten. Da Taz2 sowohl an AD1 als auch an AD2 binden kann, während Mdm2 nur AD1 bindet, ist die p53-TAD ein ideales System, um zu untersuchen, wie ein einzelnes flexibles Proteinstück Partner mit entgegengesetzten Effekten jonglieren kann.
Beobachtung einzelner Moleküle beim Partnerwechsel
Um diesen schnellen Partnerwechsel zu verfolgen, verwendeten die Forscher dreifarbige Einzelmolekül-FRET, eine Fluoreszenztechnik, die winzige Distanzänderungen innerhalb und zwischen Proteinen misst. Sie befestigten verschiedenfarbige Farbstoffe an den Enden der p53-TAD und an Taz2, während Mdm2 unlabeled blieb. Durch das Immobilisieren der TAD auf einer Glasoberfläche und das kontrollierte Zufügen der Partner konnten sie einzelne p53-Moleküle beobachten, während diese Taz2 und Mdm2 über die Zeit banden, freigesetzt wurden und wieder banden. Änderungen im Fluoreszenzmuster zeigten, ob die TAD ungebunden war, nur an Taz2 gebunden, nur an Mdm2 gebunden oder kurzzeitig an beide gleichzeitig in einem drei-Proteine-Komplex (ternärer Komplex) gebunden war.
Zwei Wege, Partner zu tauschen
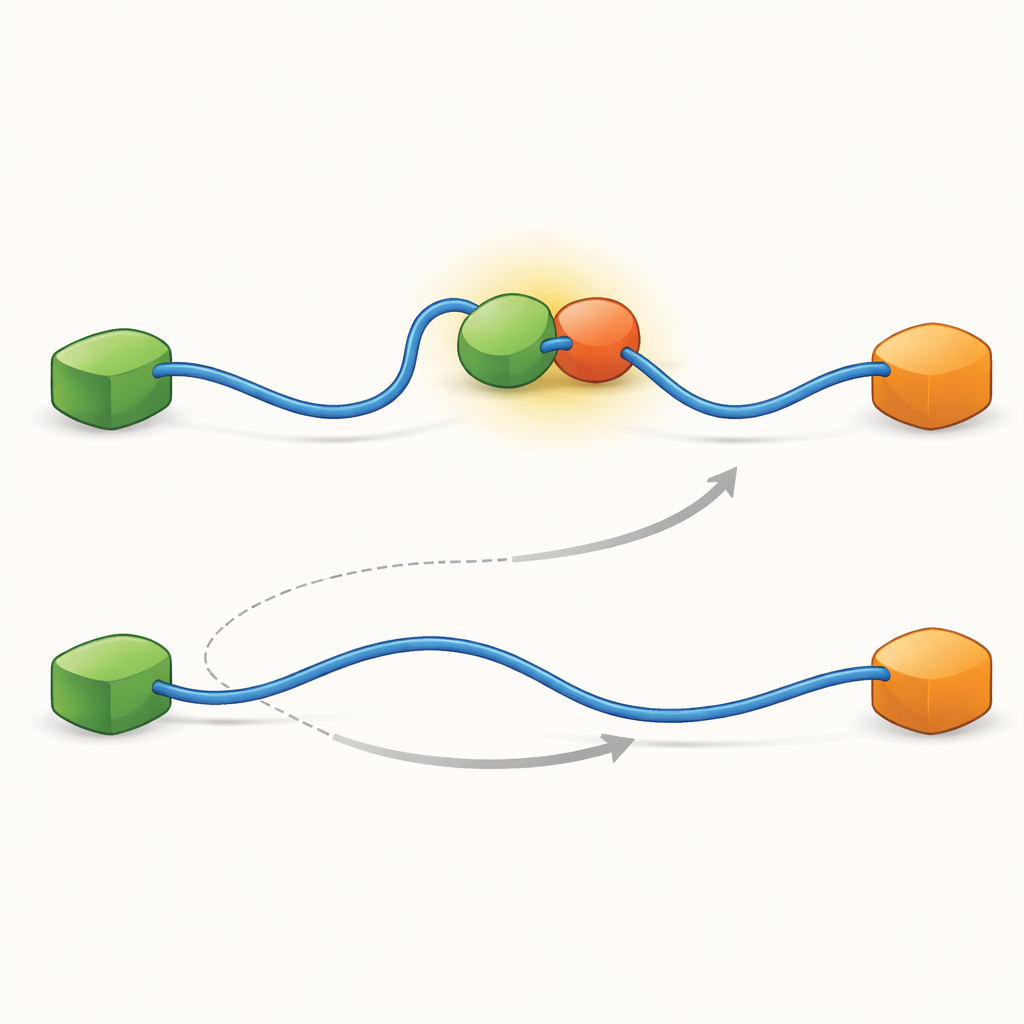
Die Aufzeichnungen zeigten, dass TAD Partner auf zwei qualitativ unterschiedliche Arten austauschen kann. Im konkurrierenden Pfad lässt die TAD zunächst einen Partner vollständig los, wird frei und flexibel und bindet erst danach den anderen Partner. Im allosterischen Pfad löst sich die TAD nicht vollständig. Stattdessen bildet sie einen kurzlebigen ternären Komplex, in dem Taz2 und Mdm2 gleichzeitig gebunden sind und jeweils einen anderen Teil der TAD greifen. Aus diesem gemeinsamen Zustand verlässt ein Partner die Bindung, während der andere verbleibt. Durch Zählen, wie lange die TAD in jedem Zustand verweilte und wie häufig Übergänge auftraten, konnten die Autoren für jeden Schritt Geschwindigkeitskonstanten ableiten und quantifizieren, wie oft jeder Pfad unter verschiedenen Bedingungen genutzt wird.
Verborgene Zwischenstufen prägen das Netzwerk
Um herauszufinden, was diese beiden Pfade ermöglicht, trieb das Team ihre Technik auf sehr hohe zeitliche Auflösung und analysierte Photonenankunftsereignisse einzeln. Sie fokussierten sich auf den Akt der TAD-Bindung an Taz2 und stellten fest, dass dieser Prozess selbst zwei unterschiedliche Arten von „Übergangspfaden“ folgt — kurzlebige Zwischenstufen zwischen ungebundenem und vollständig gebundenem Zustand. In einem Typ hält Taz2 stark an AD2, während AD1 frei bleibt; im anderen bindet Taz2 stärker an AD1, während AD2 lockerer ist. Das AD2-gebundene Zwischenstadium erlaubt es natürlich, dass Mdm2 an AD1 andockt und den ternären Komplex bildet, was den allosterischen Pfad antreibt. Dagegen blockiert das AD1-gebundene Zwischenstadium Mdm2 und zwingt das System, den konkurrenzbasierten Weg zu nutzen, bei dem die TAD sich vollständig lösen muss, bevor Mdm2 binden kann. Somit bestimmen die mikroskopischen, flüchtigen Formen, die die TAD während der Bindung annimmt, letztlich, welcher großskalige Austauschpfad dominiert.
Schnelles Umschalten für Entscheidungen über Leben oder Tod
Wenn man alle Zahlen zusammenführt, zeigen die Autoren, dass der ternäre, allosterische Pfad den Großteil des Verkehrs zwischen Mdm2-gebundenen und Taz2-gebundenen Zuständen tragen kann, besonders wenn Mdm2 reichlich vorhanden ist, wie es in Zellen der Fall ist. Dieser Weg erlaubt es p53, schnell vom durch Mdm2 gehemmtem Zustand in den durch Taz2 aktivierten Zustand umzuschalten, wenn sich die Bedingungen ändern, und gleichzeitig übermäßiges Wiederanbinden von Mdm2 zu begrenzen. Chemische Modifikationen der TAD, die die Mdm2-Bindung schwächen, können das Gleichgewicht wieder zugunsten des konkurrierenden Pfades verschieben und bieten eine zusätzliche Kontrollschicht. Einfach ausgedrückt zeigt die Studie, dass ein flexibles Proteinsegment seine Formwandlungsfähigkeit nutzt, um einen kleinen, temporären dreiseitigen Händedruck zu formen, der das Wechseln eines Partners beschleunigt. Dieser Mechanismus könnte allgemein dafür sorgen, dass ungeordnete Proteinregionen komplexe Signalnetzwerke steuern und Zellen ermöglichen, schnell auf Stress zu reagieren.
Zitation: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Schlüsselwörter: intrinsisch ungeordnete Proteine, p53-Regulation, Protein-Bindungsnetzwerke, allosterisches Umschalten, Einzelmolekül-FRET