Clear Sky Science · pl
Dynamic control of IDP interaction network via diverse binding pathways
Jak komórki podejmują szybkie decyzje
W naszych komórkach białka nieustannie wybierają partnerów, tworząc i zrywając połączenia w ułamkach sekundy. Te wybory mogą przesądzać, czy komórka rośnie, naprawia uszkodzenia, czy ulega samozniszczeniu. W badaniu tym przyjrzano się, jak jeden szczególnie istotny fragment białka supresorowego p53 szybko przełącza się między dwoma przeciwstawnymi partnerami: jednym wyłączającym aktywność p53, a drugim wspierającym jej włączenie. Obserwując pojedyncze cząsteczki w działaniu, autorzy pokazują, że to przełączanie nie przebiega jedną prostą drogą, lecz wykorzystuje dwie odrębne ścieżki, co daje komórkom szybszy i bardziej elastyczny sposób reagowania na stres.
Centrum białka zmieniającego kształt
Wiele białek w komórkach ma giętkie, nieuporządkowane regiony zwane białkami wewnętrznie nieuporządkowanymi (IDP). W przeciwieństwie do sztywnych, dobrze sfałdowanych białek, IDP mogą dopasowywać się do wielu różnych partnerów, działając jako centra w zatłoczonych sieciach molekularnych. Zespół skupił się na domenie transaktywacyjnej (TAD) na początku p53, klasycznym fragmencie IDP, który wykorzystuje dwa małe regiony, nazwane AD1 i AD2, do interakcji z wieloma regulatorami. Jeden partner, Mdm2, promuje rozkład p53 i utrzymuje jego poziom niski, gdy komórka jest zdrowa. Inny partner, Taz2, jest elementem dużego kompleksu koaktywacyjnego, który pomaga p53 włączać geny odpowiedzi na stres. Ponieważ Taz2 może wiązać zarówno AD1, jak i AD2, podczas gdy Mdm2 wiąże tylko AD1, TAD p53 jest idealnym układem do badania, jak pojedynczy elastyczny fragment białka potrafi żonglować partnerami o przeciwnych efektach.
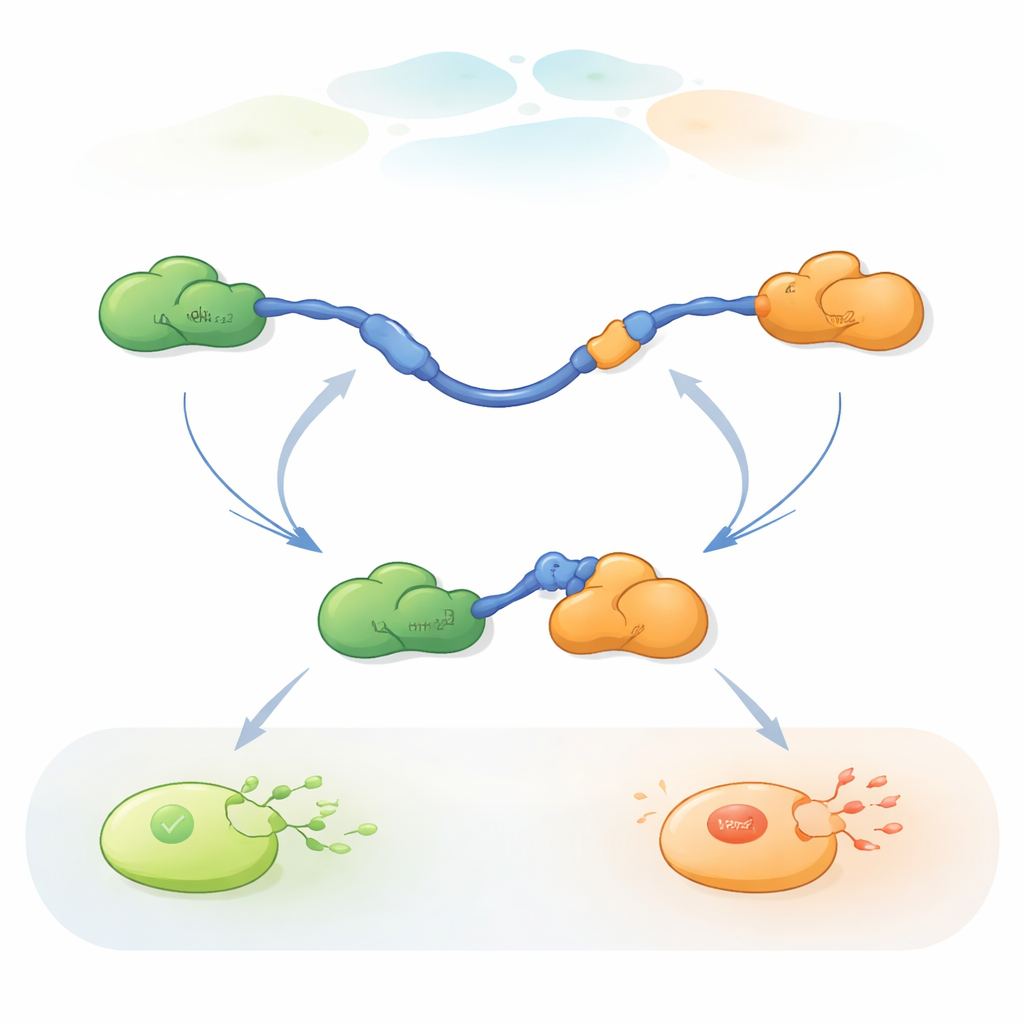
Obserwowanie pojedynczych cząsteczek wymieniających partnerów
Aby śledzić te szybkie zamiany partnerów, badacze zastosowali trójkolorowy FRET pojedynczych cząsteczek, metodę fluorescencyjną mierzącą drobne zmiany odległości wewnątrz i między białkami. Przyłączyli barwniki o różnych kolorach do końców TAD p53 i do Taz2, pozostawiając Mdm2 nieoznakowanym. Immobilizując TAD na powierzchni szklanej i dodając partnerów w kontrolowanych stężeniach, mogli obserwować pojedyncze cząsteczki p53 podczas wiązania, uwalniania i ponownego wiązania Taz2 i Mdm2 w czasie. Zmiany w wzorze fluorescencji ujawniały, czy TAD był niewiązany, związany wyłącznie z Taz2, związany wyłącznie z Mdm2, czy krótkotrwale związany z oboma jednocześnie w trzybiałkowym (ternarnym) kompleksie.
Dwa sposoby wymiany partnerów
Rejestracje pokazały, że TAD może wymieniać partnerów na dwa jakościowo różne sposoby. W ścieżce konkurencyjnej TAD najpierw całkowicie rozluźnia więź z jednym partnerem, stając się wolny i giętki, i dopiero wtedy wiąże drugiego partnera. W ścieżce allosterycznej TAD nie odłącza się w pełni. Zamiast tego tworzy krótkotrwały kompleks ternarny, w którym Taz2 i Mdm2 są związane jednocześnie, każdy chwytając inną część TAD. Z tego wspólnego stanu jeden partner odchodzi, a drugi pozostaje. Licząc, jak długo TAD pozostawał w każdym stanie i jak często występowały przejścia, autorzy mogli wyodrębnić stałe szybkości dla każdego kroku i określić, jak często każda ścieżka jest używana w różnych warunkach.
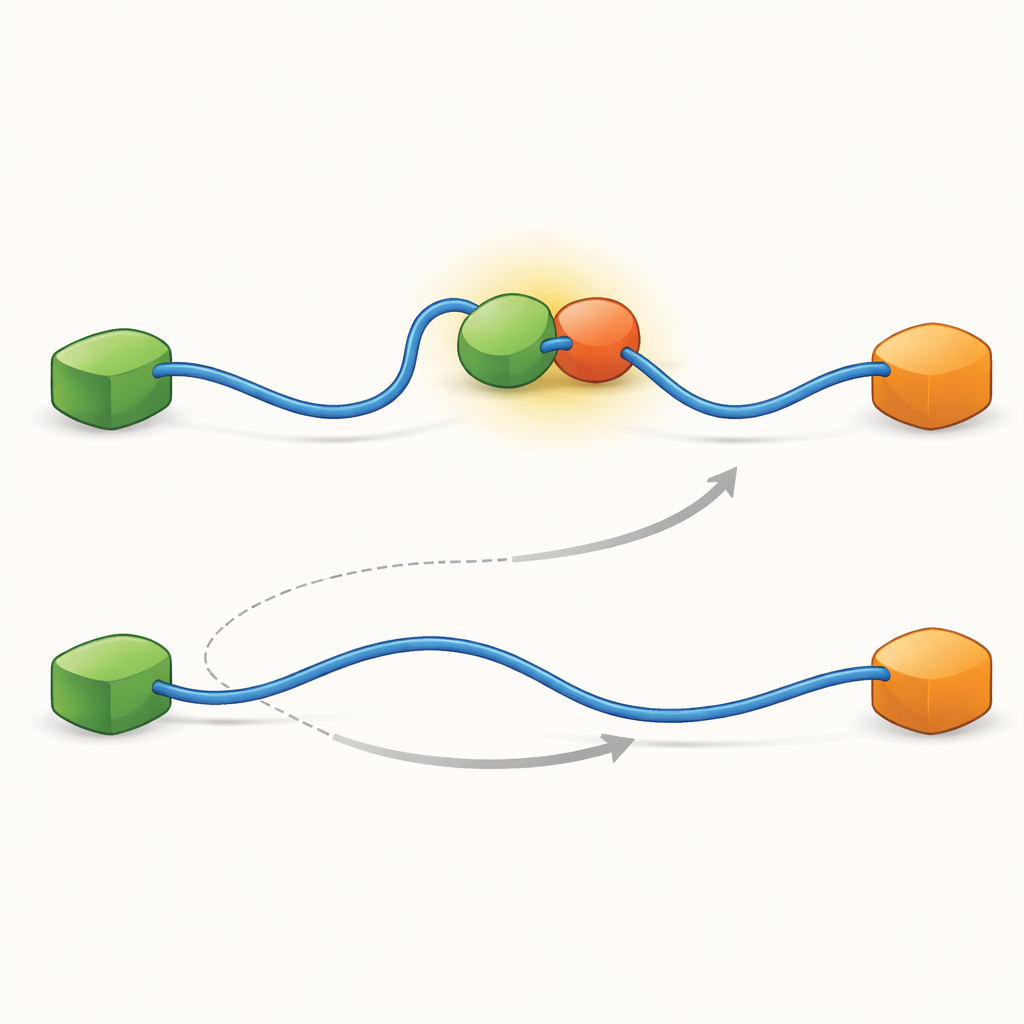
Ukryte etapy pośrednie kształtują sieć
Aby odkryć, co umożliwia istnienie tych dwóch ścieżek, zespół zwiększył rozdzielczość czasową swojej techniki i analizował zdarzenia przybycia fotonów jedno po drugim. Skoncentrowali się na akcie wiązania TAD z Taz2 i stwierdzili, że sam ten proces przebiega dwoma odrębnymi rodzajami „ścieżek przejściowych” — krótkimi etapami pośrednimi między stanem niewiązanym a w pełni związanym. W jednym typie Taz2 mocno trzyma AD2, podczas gdy AD1 pozostaje wolne; w drugim Taz2 angażuje AD1 silniej, a AD2 jest luźniejszy. Pośredni stan z AD2 związanym naturalnie pozwala Mdm2 dosunąć się do AD1 i utworzyć kompleks ternarny, zasilając ścieżkę allosteryczną. Natomiast pośredni stan z AD1 związanym blokuje Mdm2, zmuszając układ do korzystania ze ścieżki konkurencyjnej, w której TAD musi się całkowicie oderwać, zanim Mdm2 będzie mógł się związać. Tak więc mikroskopijne, krótkotrwałe kształty, które przyjmuje TAD podczas wiązania, ostatecznie decydują, która makroskopowa ścieżka wymiany przeważa.
Szybkie przełączanie dla wyborów życia lub śmierci
Składając wszystkie liczby razem, autorzy pokazują, że kompleks ternarny i związana z nim ścieżka allosteryczna mogą przenieść większość przepływu pomiędzy stanami związanymi przez Mdm2 i Taz2, szczególnie gdy Mdm2 jest obfite, jak ma to miejsce w komórkach. Ta droga pozwala p53 szybko przestawić się z bycia kontrolowanym przez Mdm2 na aktywację przez Taz2, gdy warunki się zmieniają, jednocześnie ograniczając nadmierne ponowne wiązanie Mdm2. Modyfikacje chemiczne TAD osłabiające wiązanie Mdm2 mogą przesunąć równowagę z powrotem ku ścieżce konkurencyjnej, oferując dodatkową warstwę kontroli. W prostych słowach, badanie ujawnia, że elastyczny fragment białka wykorzystuje swoją zdolność do zmiany kształtu, by zbudować małe, tymczasowe trójstronne uściskanie dłoni, które przyspiesza wymianę jednego partnera na drugiego. Mechanizm ten może być ogólnym sposobem, w jaki nieuporządkowane regiony białek zarządzają złożonymi sieciami sygnalizacyjnymi i pomagają komórkom szybko reagować na stres.
Cytowanie: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Słowa kluczowe: białka wewnętrznie nieuporządkowane, regulacja p53, sieci wiązań białkowych, przełączanie allosteryczne, FRET pojedynczych cząsteczek