Clear Sky Science · en
Dynamic control of IDP interaction network via diverse binding pathways
How Cells Make Fast Decisions
Inside our cells, proteins are constantly choosing partners, forming and breaking connections in fractions of a second. These choices can decide whether a cell grows, repairs damage, or self-destructs. This study looks at how one especially important protein segment from the tumor suppressor p53 rapidly switches between two opposing partners: one that turns p53 activity off and another that helps turn it on. By watching single molecules in action, the authors show that this switching does not follow a single simple route, but instead uses two distinct pathways that give cells a faster and more flexible way to respond to stress.
A Shape-Shifting Protein Hub
Many proteins in our cells have floppy, unstructured regions known as intrinsically disordered proteins (IDPs). Unlike rigid, well-folded proteins, IDPs can mold themselves to fit many different partners, acting as hubs in crowded molecular networks. The team focused on the transactivation domain (TAD) at the beginning of p53, a classic IDP segment that uses two small regions, called AD1 and AD2, to interact with many regulators. One partner, Mdm2, promotes p53 breakdown and keeps its levels low when the cell is healthy. Another partner, Taz2, is part of a large coactivator complex that helps p53 turn on stress-response genes. Because Taz2 can bind both AD1 and AD2 while Mdm2 binds only AD1, p53 TAD is an ideal system to ask how a single flexible piece of protein can juggle partners with opposite effects.
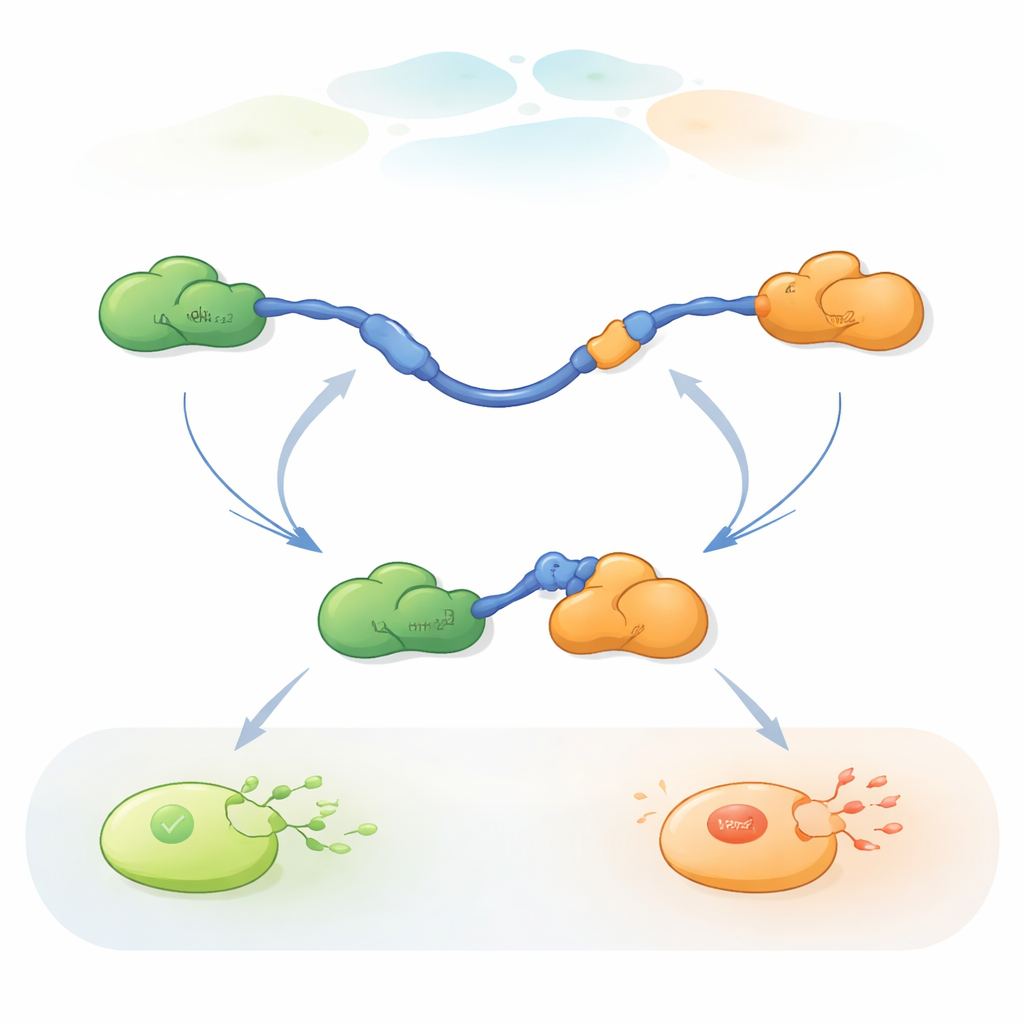
Watching Single Molecules Switch Partners
To follow these rapid partner swaps, the researchers used three-color single-molecule FRET, a fluorescence method that measures tiny distance changes within and between proteins. They attached different colored dyes to the ends of p53 TAD and to Taz2, while leaving Mdm2 unlabeled. By immobilizing TAD on a glass surface and adding the partners in controlled amounts, they could watch individual p53 molecules as they bound, released, and re-bound Taz2 and Mdm2 over time. Changes in the fluorescence pattern revealed whether TAD was unbound, bound only to Taz2, bound only to Mdm2, or briefly bound to both at once in a three-protein (ternary) complex.
Two Ways to Trade Partners
The recordings showed that TAD can exchange partners in two qualitatively different ways. In the competitive pathway, TAD first lets go of one partner completely, becoming free and flexible, and only then binds the other partner. In the allosteric pathway, TAD does not fully detach. Instead, it forms a short-lived ternary complex where Taz2 and Mdm2 are both bound at the same time, each gripping a different part of TAD. From this joint state, one partner leaves and the other remains. By counting how long TAD stayed in each state and how often transitions occurred, the authors could extract rate constants for every step and quantify how frequently each pathway is used under different conditions.
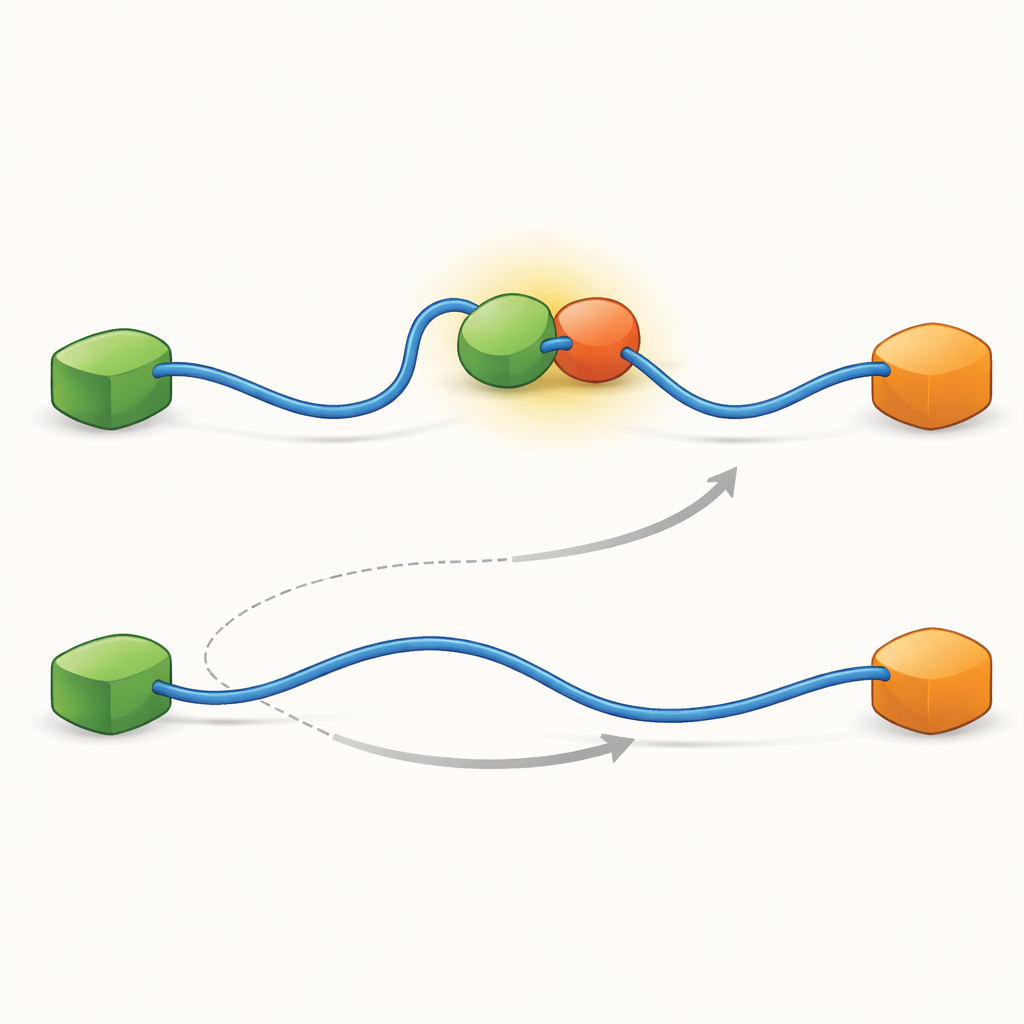
Hidden Intermediate Steps Shape the Network
To uncover what makes these two pathways possible, the team pushed their technique to very high time resolution and analyzed photon arrival events one by one. They focused on the act of TAD binding to Taz2 and found that this process itself follows two distinct kinds of “transition paths”—brief intermediate stages between unbound and fully bound states. In one type, Taz2 holds strongly to AD2 while AD1 remains free; in the other, Taz2 engages AD1 more strongly while AD2 is looser. The AD2-bound intermediate naturally allows Mdm2 to slip onto AD1 and create the ternary complex, feeding the allosteric pathway. By contrast, the AD1-bound intermediate blocks Mdm2, forcing the system to use the competitive route where TAD must fully detach before Mdm2 can bind. Thus, the microscopic, fleeting shapes that TAD adopts during binding end up dictating which large-scale exchange pathway dominates.
Fast Switching for Life-or-Death Choices
Putting all the numbers together, the authors show that the ternary-complex, allosteric pathway can carry most of the traffic between Mdm2-bound and Taz2-bound states, especially when Mdm2 is abundant, as it is in cells. This route lets p53 quickly switch from being held in check by Mdm2 to being activated by Taz2 when conditions change, while still limiting excess rebinding of Mdm2. Chemical modifications to TAD that weaken Mdm2 binding can shift the balance back toward the competitive pathway, offering another layer of control. In simple terms, the study reveals that a flexible protein segment uses its shape-shifting nature to build a small, temporary three-way handshake that speeds up trading one partner for another. This mechanism may be a general way that disordered protein regions manage complex signaling networks and help cells respond rapidly to stress.
Citation: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Keywords: intrinsically disordered proteins, p53 regulation, protein binding networks, allosteric switching, single-molecule FRET