Clear Sky Science · ar
التحكم الديناميكي بشبكة تفاعلات البروتينات غير المرتبة عبر مسارات ارتباط متنوعة
كيف تتخذ الخلايا قرارات سريعة
داخل خلايانا، تختار البروتينات شركاءها باستمرار، مكوِّنة ومفكِّكة روابط في أجزاء من الثانية. يمكن أن تحدد هذه الاختيارات ما إذا كانت الخلية ستنمو أو تصلّح الضرر أو تنهار ذاتيًا. تبحث هذه الدراسة في كيفية تَبدّل مقطع بروتيني مهم من مثبط الورم p53 بسرعة بين شريكين متعارضين: أحدهما يوقف نشاط p53 والآخر يساعد في تفعيله. بمراقبة جزيئات مفردة أثناء العمل، يبيّن المؤلفون أن هذا التبديل لا يتبع مسارًا وحيدًا بسيطًا، بل يستخدم مسارين متميزين يمنحان الخلايا طريقة أسرع وأكثر مرونة للاستجابة للإجهاد.
مركز بروتيني متبدل الشكل
تحوي العديد من البروتينات في خلايانا مناطق رخوة وغير منظمة تُعرف بالبروتينات غير المرتبة جوهريًا (IDPs). على عكس البروتينات الصلبة والمطوية جيدًا، يمكن لهذه المناطق أن تشكِّل نفسها لتتناسب مع شركاء مختلفين، فتلعب دور محاور في شبكات جزيئية مزدحمة. ركّز الفريق على نطاق التشغيل (TAD) في بداية p53، وهو مقطع نموذجي من IDP يستخدم منطقتين صغيرتين تُدعيان AD1 وAD2 للتفاعل مع العديد من المنظمين. أحد الشركاء، Mdm2، يعزز تحلل p53 ويحافظ على مستوياته منخفضة عندما تكون الخلية سليمة. الشريك الآخر، Taz2، جزء من مركب كبير مساعد للتفعيل يساعد p53 في تشغيل جينات استجابة الإجهاد. لأن Taz2 يمكنه الارتباط بكل من AD1 وAD2 بينما يرتبط Mdm2 بـAD1 فقط، فإن نطاق TAD في p53 يشكل نظامًا مثاليًا لطرح سؤال كيف يمكن لقطعة بروتينية مرنة واحدة أن توازن بين شركاء لهم آثار متعاكسة.
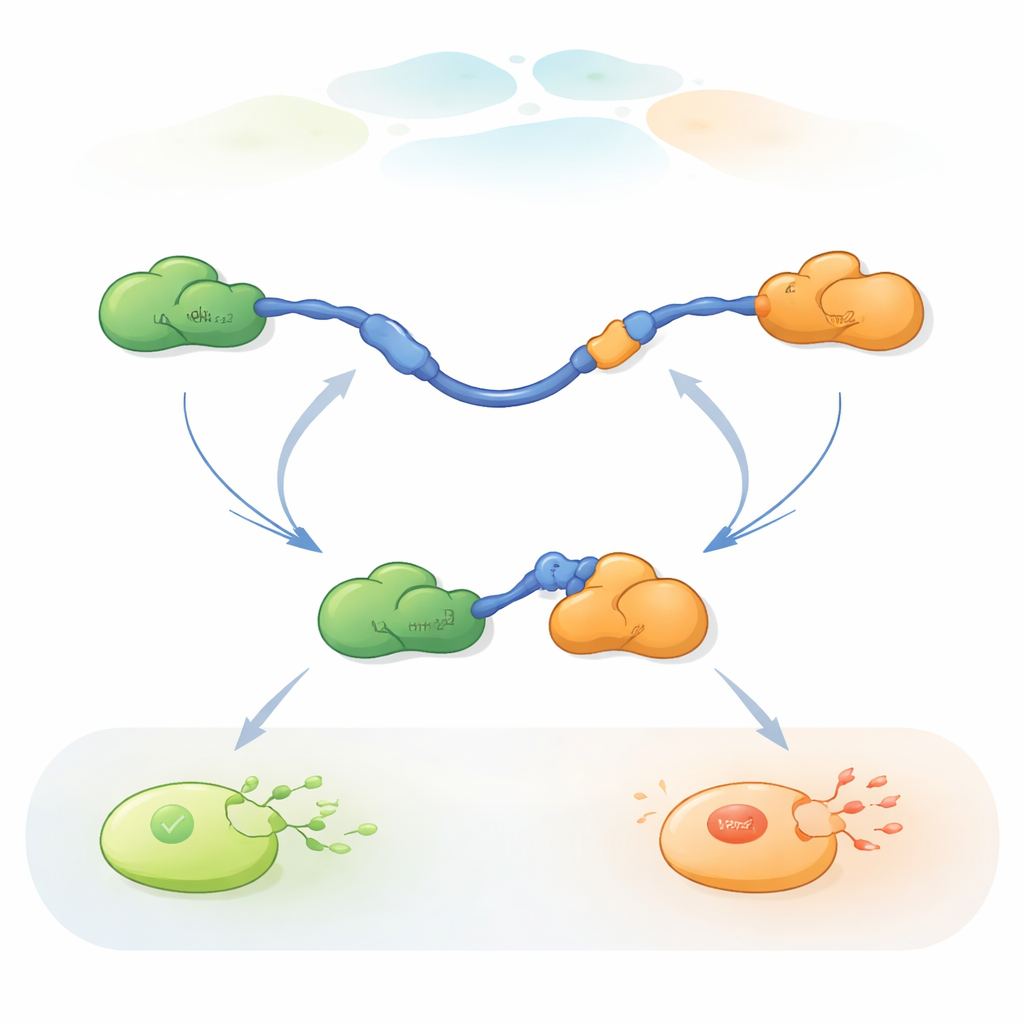
مراقبة تبديل الشركاء على مستوى جزيء واحد
لمتابعة هذه التبادلات السريعة للشركاء، استخدم الباحثون تقنية الفِرْت ثلاثية الألوان على مستوى الجزيء الواحد، وهي طريقة تألق تقيس تغيّرات طفيفة في المسافات داخل البروتينات وبينها. وضعوا صبغات بألوان مختلفة على أطراف نطاق TAD من p53 وعلى Taz2، بينما تركوا Mdm2 غير معنَّم. من خلال تثبيت TAD على سطح زجاجي وإضافة الشركاء بكميات مضبوطة، تمكنوا من مشاهدة جزيئات p53 المفردة أثناء ارتباطها وفك ارتباطها وإعادة ارتباطها بـTaz2 وMdm2 مع مرور الوقت. كشفت التغييرات في نمط التألق عما إذا كان TAD غير مرتبط أو مرتبطًا فقط بـTaz2 أو مرتبطًا فقط بـMdm2، أو مرتبطًا لفترة وجيزة بكليهما في آن واحد ضمن مركب ثلاثي البروتينات (ثلاثي).
طريقتان لتبديل الشركاء
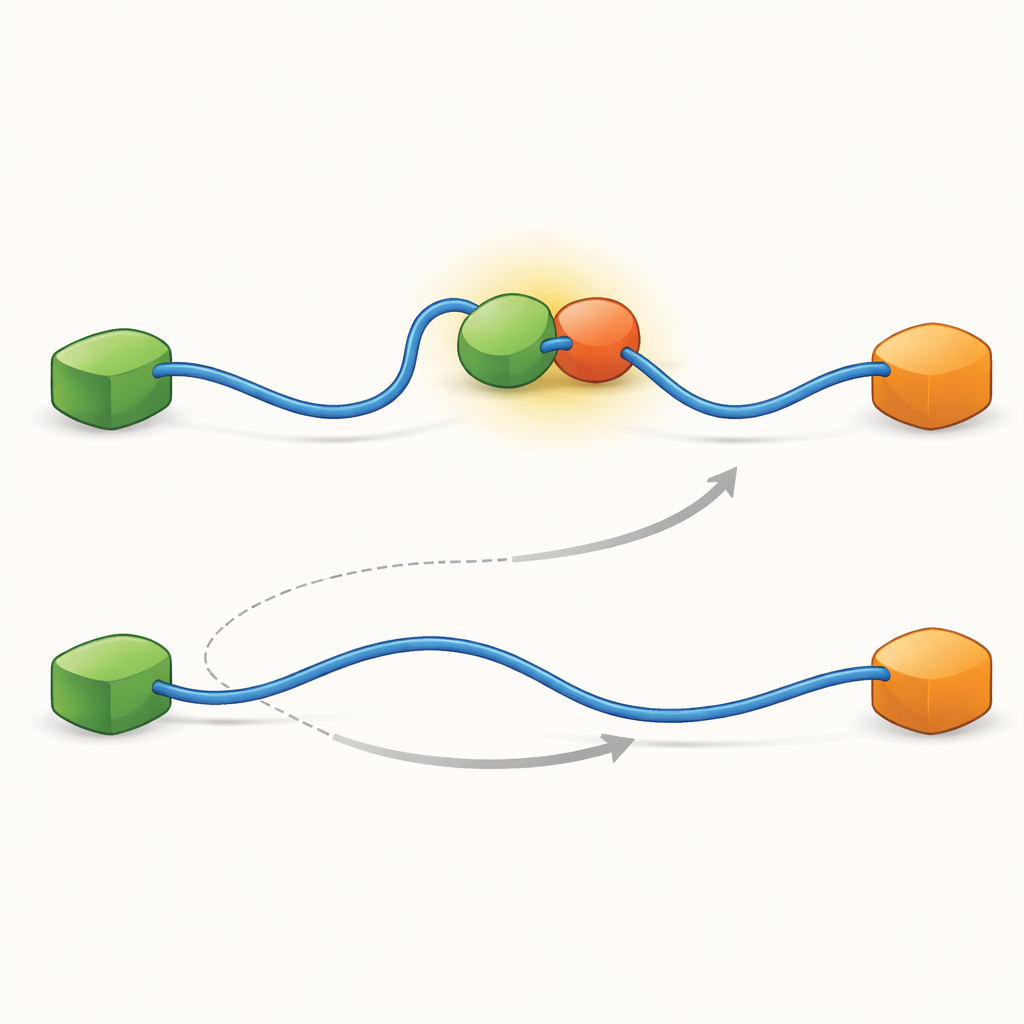
أظهرت التسجيلات أن TAD يمكنه تبديل الشركاء بطريقتين مختلفتين نوعيًا. في المسار التنافسي، يطلق TAD شريكًا واحدًا تمامًا أولًا، فيصبح حرًا ومرنًا، ثم يرتبط بالشريك الآخر بعد ذلك فقط. في المسار التآزري، لا ينفصل TAD بالكامل؛ بدلاً من ذلك يشكل مركبًا ثلاثيًّا قصير العمر حيث يرتبط كل من Taz2 وMdm2 في الوقت نفسه، كلٌ يمسك جزءًا مختلفًا من TAD. من هذه الحالة المشتركة يغادر أحد الشركاء بينما يبقى الآخر. من خلال عدّ المدد الزمنية التي قضّاها TAD في كل حالة وعدد التحولات، استطاع المؤلفون استخلاص ثوابت السرعة لكل خطوة وقياس مدى تكرار استعمال كل مسار تحت ظروف مختلفة.
خطوات وسيطة خفية تشكّل الشبكة
لكشف ما يجعل هذين المسارين ممكنين، دفع الفريق تقنيتهم إلى دقة زمنية عالية جدًا وحللوا أحداث وصول الفوتونات واحدًا فواحد. ركزوا على فعل ارتباط TAD بـTaz2 ووجدوا أن هذه العملية نفسها تتبع نوعين متميزين من «مسارات الانتقال» — مراحل وسيطة وجيزة بين الحالات غير المرتبطة والمرتبطة بالكامل. في نوع واحد، يمسك Taz2 بقوة بـAD2 بينما يبقى AD1 حرًا؛ في النوع الآخر، يتفاعل Taz2 مع AD1 بقوة أكبر بينما يكون AD2 أكثر ارتخاءً. الوسيط المرتبط بـAD2 يسمح بطبيعة الحال لـMdm2 بالتقاط AD1 وتشكيل المركب الثلاثي، مما يغذي المسار التآزري. بالمقابل، الوسيط المرتبط بـAD1 يمنع Mdm2، مما يضطر النظام لاستخدام الطريق التنافسية حيث يجب على TAD الانفصال كليًا قبل أن يستطيع Mdm2 الارتباط. هكذا، فإن الأشكال الميكروسكوبية العابرة التي يتبناها TAD أثناء الارتباط هي التي تحدد أي مسار تبادل واسع النطاق يسود.
تبديل سريع لقرارات مصيرية
بجمع كل الأرقام، يُظهر المؤلفون أن المسار التآزري عبر المركب الثلاثي يمكن أن يحمل معظم الحركة بين الحالات المرتبطة بـMdm2 وتلك المرتبطة بـTaz2، لا سيما عندما يكون Mdm2 وافرًا كما هو الحال في الخلايا. تتيح هذه الطريق لـp53 أن يتحول بسرعة من حالة محجوزة بواسطة Mdm2 إلى حالة مفعَّلة بواسطة Taz2 عند تغيّر الظروف، مع الحد من الارتباط الزائد المتكرر لـMdm2. التعديلات الكيميائية على TAD التي تضعف ارتباط Mdm2 يمكن أن تُرجِح التوازن نحو المسار التنافسي، موفرة طبقة تحكّم إضافية. بعبارة بسيطة، تكشف الدراسة أن مقطعًا بروتينيًا مرنًا يستخدم طبيعته المتغيرة الشكل لبناء مصافحة ثلاثية مؤقتة تُسرّع تبادل الشريك. قد يكون هذا الميكانيزم وسيلة عامة تستخدمها المناطق البروتينية غير المرتبة لإدارة شبكات الإشارة المعقدة ومساعدة الخلايا على الاستجابة السريعة للإجهاد.
الاستشهاد: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
الكلمات المفتاحية: البروتينات غير المرتبة جوهريًا, تنظيم p53, شبكات ارتباط البروتينات, التحويل التآزري, الفّرت أحادي الجزئيات