Clear Sky Science · sv
Dynamisk kontroll av IDP-interaktionsnätverk via olika bindningsvägar
Hur celler fattar snabba beslut
Inne i våra celler väljer proteiner ständigt partner, bildar och bryter kopplingar på bråkdelar av en sekund. Dessa val kan avgöra om en cell växer, reparerar skador eller självdestruerar. Denna studie undersöker hur ett särskilt viktigt proteinsekt från tumörsupressorn p53 snabbt växlar mellan två motsatta partners: en som stänger av p53:s aktivitet och en annan som hjälper till att slå på den. Genom att följa enstaka molekyler i arbete visar författarna att denna växling inte följer en enda enkel väg, utan istället använder två distinkta banor som ger celler ett snabbare och mer flexibelt sätt att reagera på stress.
En formskiftande proteinknutpunkt
Många proteiner i våra celler har lösa, oordnade regioner som kallas intrinsikt oordnade proteiner (IDP). Till skillnad från styva, välveckade proteiner kan IDP:er formas om för att passa många olika partners och fungera som nav i täta molekylära nätverk. Gruppen fokuserade på transaktiveringsdomänen (TAD) i början av p53, ett klassiskt IDP-segment som använder två små regioner, kallade AD1 och AD2, för att interagera med många regulatorer. En partner, Mdm2, främjar p53-nedbrytning och håller nivåerna låga när cellen är frisk. En annan partner, Taz2, är del av ett stort koaktivatorkomplex som hjälper p53 att slå på genprogram för stressrespons. Eftersom Taz2 kan binda både AD1 och AD2 medan Mdm2 bara binder AD1, är p53 TAD ett idealiskt system för att fråga hur ett enda flexibelt proteinknäpp kan jonglera partners med motsatta effekter.
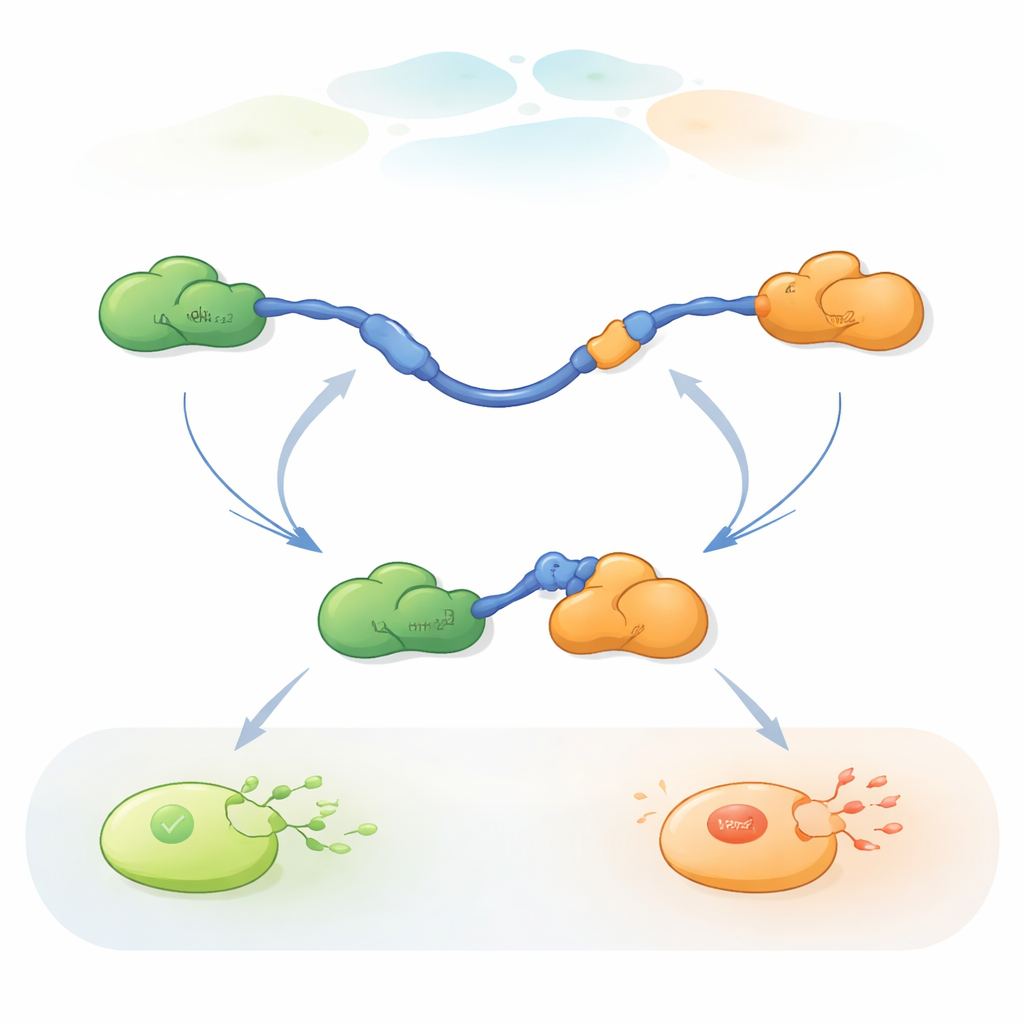
Att se enstaka molekyler byta partner
För att följa dessa snabba partnerbyten använde forskarna trefärgad single-molecule FRET, en fluorescensmetod som mäter små avståndsändringar inom och mellan proteiner. De fäste olika färgade färgämnen på ändarna av p53 TAD och på Taz2, medan Mdm2 lämnades omärkt. Genom att immobilisera TAD på en glasyta och tillsätta partnerna i kontrollerade mängder kunde de se enskilda p53-molekyler när de bundit, släppt och återbundet Taz2 och Mdm2 över tid. Förändringar i fluorescensmönstret avslöjade om TAD var obundet, bundet endast till Taz2, bundet endast till Mdm2 eller kortvarigt bundet till båda samtidigt i ett ternärt komplex med tre proteiner.
Två sätt att byta partner
Inspelningarna visade att TAD kan byta partner på två kvalitativt olika sätt. I den konkurrerande vägen släpper TAD först en partner helt, blir fri och flexibel, och binder sedan den andra partnern. I den allosteriska vägen lossnar inte TAD helt. Istället bildar det ett kortlivat ternärt komplex där Taz2 och Mdm2 båda är bundna samtidigt, var och en greppar en annan del av TAD. Från detta gemensamma tillstånd lämnar en partner och den andra blir kvar. Genom att räkna hur länge TAD stannade i varje tillstånd och hur ofta övergångar inträffade kunde författarna dra ut hastighetskonstanter för varje steg och kvantifiera hur ofta varje väg används under olika förhållanden.
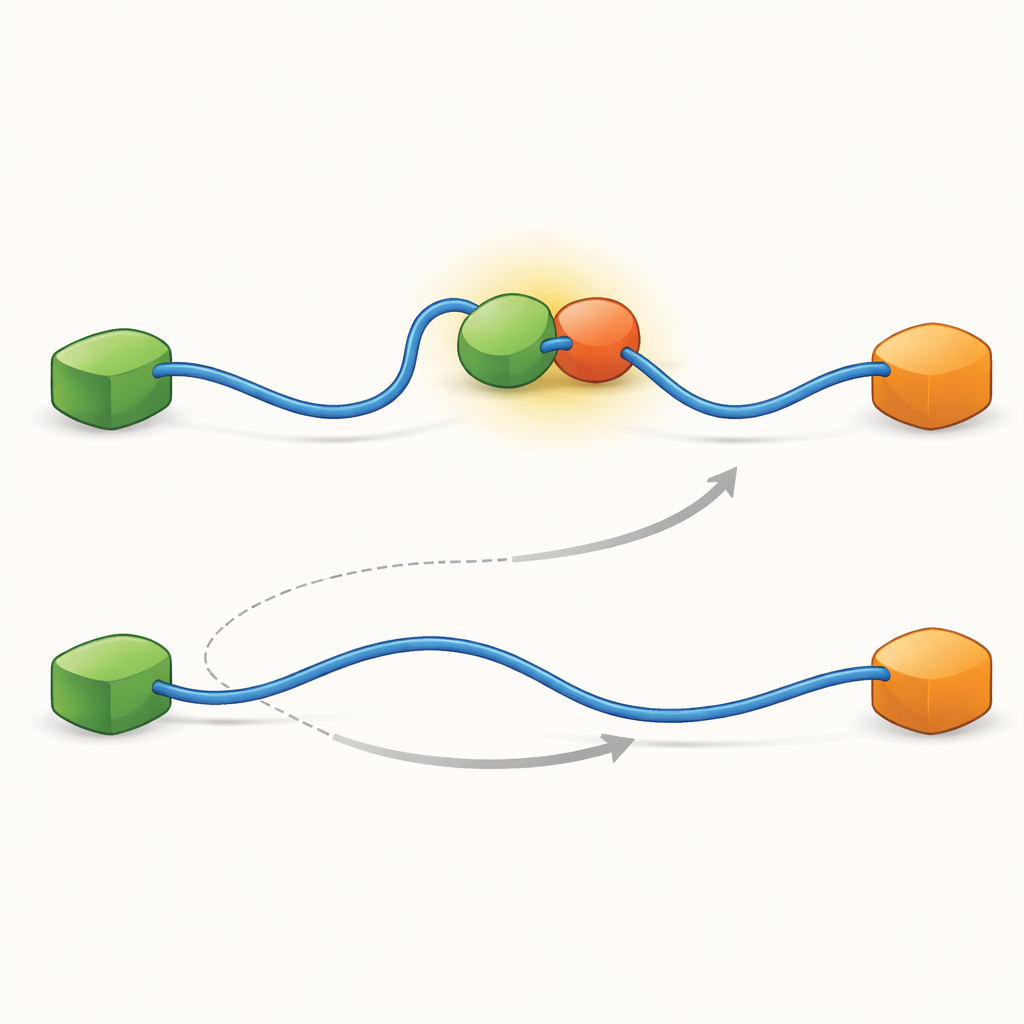
Dolda intermediära steg formar nätverket
För att avslöja vad som möjliggör dessa två vägar pressade teamet sin teknik till mycket hög tidsupplösning och analyserade fotonankomsthändelser en och en. De fokuserade på själva bindningen av TAD till Taz2 och fann att denna process i sig följer två distinkta typer av ”övergångsvägar” — korta intermediära stadier mellan obundet och fullt bundet tillstånd. I den ena typen håller Taz2 starkt i AD2 medan AD1 förblir fri; i den andra engagerar Taz2 AD1 mer kraftfullt medan AD2 är lösare. Det AD2-bundna intermediatet tillåter naturligt att Mdm2 glider på AD1 och skapar det ternära komplexet, vilket matar den allosteriska vägen. Däremot blockerar det AD1-bundna intermediatet Mdm2 och tvingar systemet att använda den konkurrerande rutten där TAD måste lossna helt innan Mdm2 kan binda. Således dikterar de mikroskopiska, flyktiga former som TAD antar under bindning vilken storskalig utbytesväg som dominerar.
Snabb växling för liv-eller-död-val
Genom att lägga ihop alla siffror visar författarna att det ternära, allosteriska förloppet kan bära större delen av trafiken mellan Mdm2-bundna och Taz2-bundna tillstånd, särskilt när Mdm2 är rikligt, som det är i celler. Denna väg låter p53 snabbt gå från att hållas i schack av Mdm2 till att aktiveras av Taz2 när förhållandena förändras, samtidigt som överdriven återbindning av Mdm2 begränsas. Kemiska modifieringar av TAD som försvagar Mdm2-bindning kan skifta balansen tillbaka mot den konkurrerande vägen, vilket ger ytterligare ett styrskikt. I enkla termer visar studien att ett flexibelt proteinsegment använder sin formskiftande natur för att bygga en liten, temporär trevägshälsning som snabbar upp utbytet av en partner mot en annan. Denna mekanism kan vara ett generellt sätt som oordnade proteinregioner hanterar komplexa signalnätverk och hjälper celler att reagera snabbt på stress.
Citering: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Nyckelord: intrinsiskt oordnade proteiner, p53-reglering, proteinbindningsnätverk, allosterisk omkoppling, single-molecule FRET