Clear Sky Science · ru
Динамическое управление сетью взаимодействий ИББ посредством различных путей связывания
Как клетки принимают быстрые решения
Внутри наших клеток белки постоянно выбирают партнёров, устанавливая и разрывая связи за доли секунды. Эти выборы могут определить, будет ли клетка расти, восстанавливать повреждения или погибать. В этом исследовании рассматривается, как один особенно важный сегмент белка-супрессора опухолей p53 быстро переключается между двумя противоположными партнёрами: одним, отключающим активность p53, и другим, способствующим её включению. Наблюдая за отдельными молекулами в действии, авторы показали, что это переключение не следует единому простому пути, а использует два разных маршрута, давая клеткам более быстрый и гибкий ответ на стресс.
Центр, меняющий форму
Многие белки в наших клетках имеют гибкие, неструктурированные участки, известные как внутренне неупорядоченные белки (ИББ). В отличие от жёстко свернутых белков, ИББ могут принимать разные формы, подстраиваясь под множество партнёров и действуя как узлы в густонаселённых молекулярных сетях. Команда сосредоточилась на трансацтивном домене (TAD) в начале p53 — классическом сегменте ИББ, который использует две небольшие области, называемые AD1 и AD2, для взаимодействия со множеством регуляторов. Один партнёр, Mdm2, способствует разрушению p53 и поддерживает его низкие уровни в здоровой клетке. Другой партнёр, Taz2, входит в состав крупного коактиваторного комплекса, помогающего p53 включать гены ответа на стресс. Поскольку Taz2 может связываться с обоими — AD1 и AD2, а Mdm2 — только с AD1, TAD p53 служит идеальной системой для изучения того, как одна гибкая часть белка может жонглировать партнёрами с противоположными эффектами.
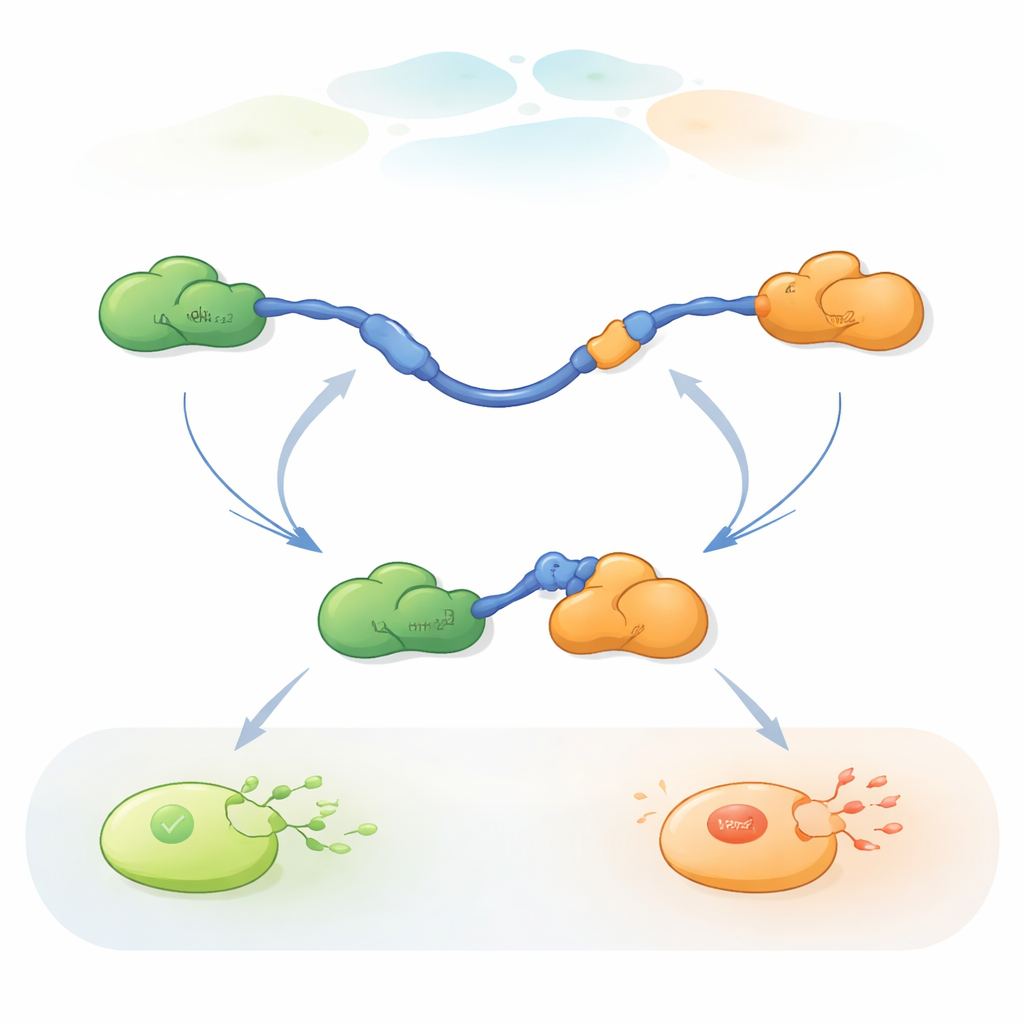
Наблюдая, как отдельные молекулы меняют партнёров
Чтобы проследить эти быстрые обмены партнёров, исследователи использовали трёхцветный одномолекулярный FRET — флуоресцентный метод, измеряющий крошечные изменения расстояний внутри и между белками. Они прикрепляли красители разных цветов к концам TAD p53 и к Taz2, оставляя Mdm2 немеченым. Фиксируя TAD на стеклянной поверхности и добавляя партнёров в контролируемых концентрациях, они могли наблюдать отдельные молекулы p53 по мере их связывания, освобождения и повторного связывания с Taz2 и Mdm2 во времени. Изменения в шаблоне флуоресценции показывали, был ли TAD несвязан, связан только с Taz2, связан только с Mdm2 или кратковременно одновременно связан с обоими в трёхбелковом (тернарном) комплексе.
Два пути обмена партнёрами
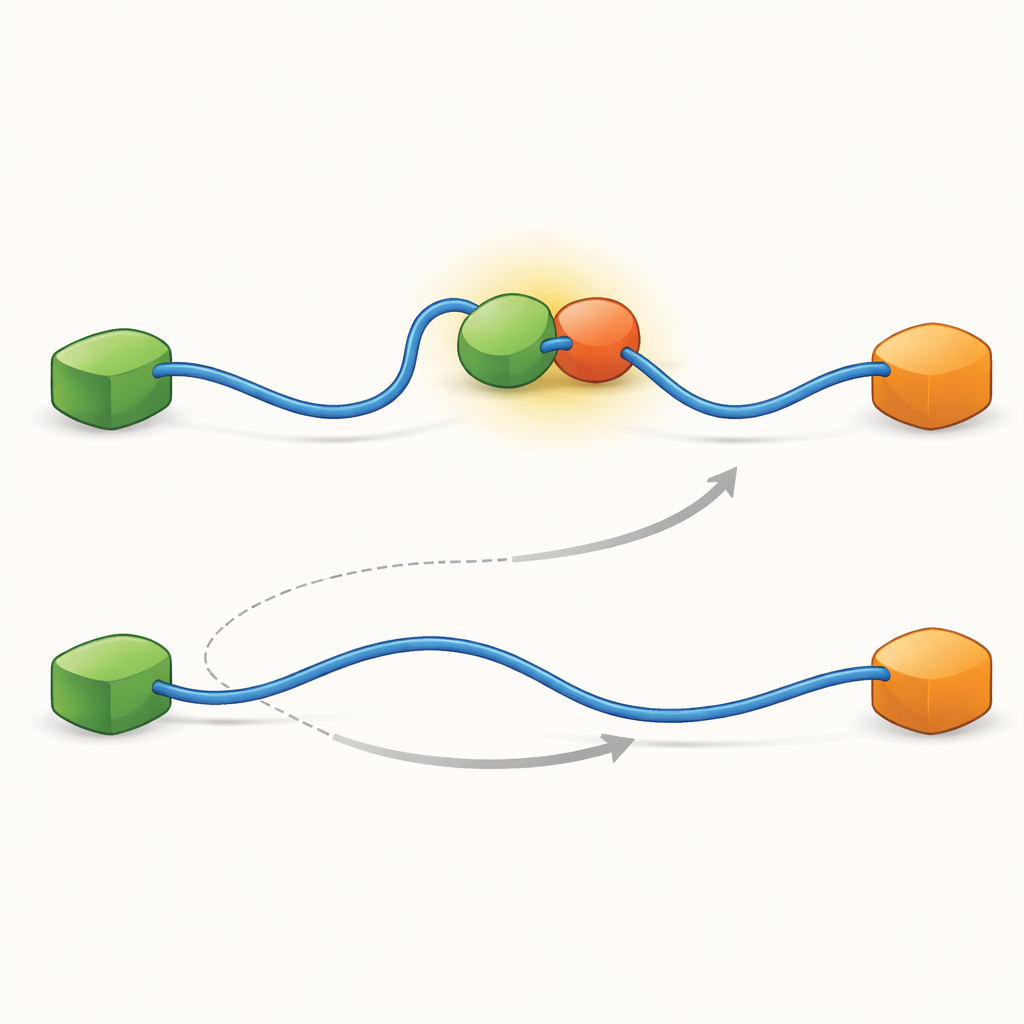
Записи показали, что TAD может обменивать партнёров двумя качественно разными способами. В конкурентном пути TAD сначала полностью отпускает одного партнёра, становясь свободным и гибким, и только затем связывается с другим партнёром. В аллостерическом пути TAD не отрывается полностью. Вместо этого он образует кратковременный тернарный комплекс, где Taz2 и Mdm2 одновременно связаны, каждый удерживая разную часть TAD. Из этого совместного состояния один партнёр уходит, а другой остаётся. Подсчитав, как долго TAD пребывал в каждом состоянии и как часто происходили переходы, авторы смогли извлечь константы скоростей для каждого шага и количественно оценить, насколько часто используется каждый путь при разных условиях.
Скрытые промежуточные этапы формируют сеть
Чтобы выяснить, что делает возможными эти два пути, команда довела методику до очень высокой временной разрешающей способности и проанализировала события прихода фотонов по одному. Они сосредоточились на акте связывания TAD с Taz2 и обнаружили, что сам этот процесс следует двум разным типам «переходных путей» — кратким промежуточным стадиям между несвязанным и полностью связанным состоянием. В одном типе Taz2 крепко удерживает AD2, тогда как AD1 остаётся свободным; в другом типе Taz2 сильнее вовлекает AD1, а AD2 более свободен. Промежуточное состояние с AD2, связанным с Taz2, естественно позволяет Mdm2 пристроиться к AD1 и образовать тернарный комплекс, обеспечивая аллостерический путь. Напротив, промежуточное состояние с AD1, связанным с Taz2, блокирует Mdm2, заставляя систему идти конкурентным путём, где TAD должен полностью отделиться прежде, чем Mdm2 сможет связаться. Таким образом микроскопические, мимолётные формы, которые принимает TAD при связывании, в конечном счёте определяют, какой крупномасштабный путь обмена доминирует.
Быстрое переключение для решений «жизнь или смерть»
Собрав все числа вместе, авторы показывают, что тернарный, аллостерический путь может нести большую часть трафика между состояниями, связанными с Mdm2 и Taz2, особенно когда Mdm2 в избытке, как это бывает в клетках. Этот путь позволяет p53 быстро переключаться от подавления Mdm2 к активации Taz2 при изменении условий, одновременно ограничивая чрезмерное повторное связывание с Mdm2. Химические модификации TAD, ослабляющие связывание с Mdm2, могут смещать баланс в сторону конкурентного пути, создавая ещё один уровень контроля. Проще говоря, исследование показывает, что гибкий сегмент белка использует свою меняющуюся форму, чтобы создать небольшое временное трёхстороннее рукопожатие, ускоряющее обмен партнёрами. Этот механизм может быть общим способом, с помощью которого неупорядоченные участки белков управляют сложными сигнальными сетями и помогают клеткам быстро реагировать на стресс.
Цитирование: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Ключевые слова: внутренне неупорядоченные белки, регуляция p53, сети связывания белков, аллостерическое переключение, одномолекулярный FRET