Clear Sky Science · it
Controllo dinamico della rete di interazioni degli IDP tramite percorsi di legame diversi
Come le cellule prendono decisioni rapide
All'interno delle nostre cellule, le proteine scelgono costantemente partner, formando e spezzando connessioni in frazioni di secondo. Queste scelte possono determinare se una cellula cresce, ripara danni o si autodistrugge. Questo studio esamina come un segmento proteico particolarmente importante del soppressore tumorale p53 passi rapidamente tra due partner opposti: uno che spegne l'attività di p53 e un altro che la attiva. Osservando molecole singole in azione, gli autori mostrano che questo cambio non segue un’unica via semplice, ma utilizza invece due percorsi distinti che conferiscono alla cellula un modo più veloce e flessibile di rispondere allo stress.
Un hub proteico che cambia forma
Molte proteine nelle nostre cellule possiedono regioni flessuose e non strutturate note come proteine intrinsecamente disordinate (IDP). A differenza delle proteine rigide e ben ripiegate, le IDP possono adattarsi per interagire con molti partner diversi, fungendo da hub in reti molecolari affollate. Il gruppo si è concentrato sul dominio di transattivazione (TAD) all'inizio di p53, un segmento IDP classico che usa due piccole regioni, chiamate AD1 e AD2, per interagire con numerosi regolatori. Un partner, Mdm2, promuove la degradazione di p53 mantenendone bassa la concentrazione quando la cellula è sana. Un altro partner, Taz2, fa parte di un grande complesso coattivatore che aiuta p53 ad attivare geni della risposta allo stress. Poiché Taz2 può legare sia AD1 sia AD2 mentre Mdm2 lega solo AD1, il TAD di p53 è un sistema ideale per chiedersi come un singolo pezzo proteico flessibile possa gestire partner con effetti opposti.
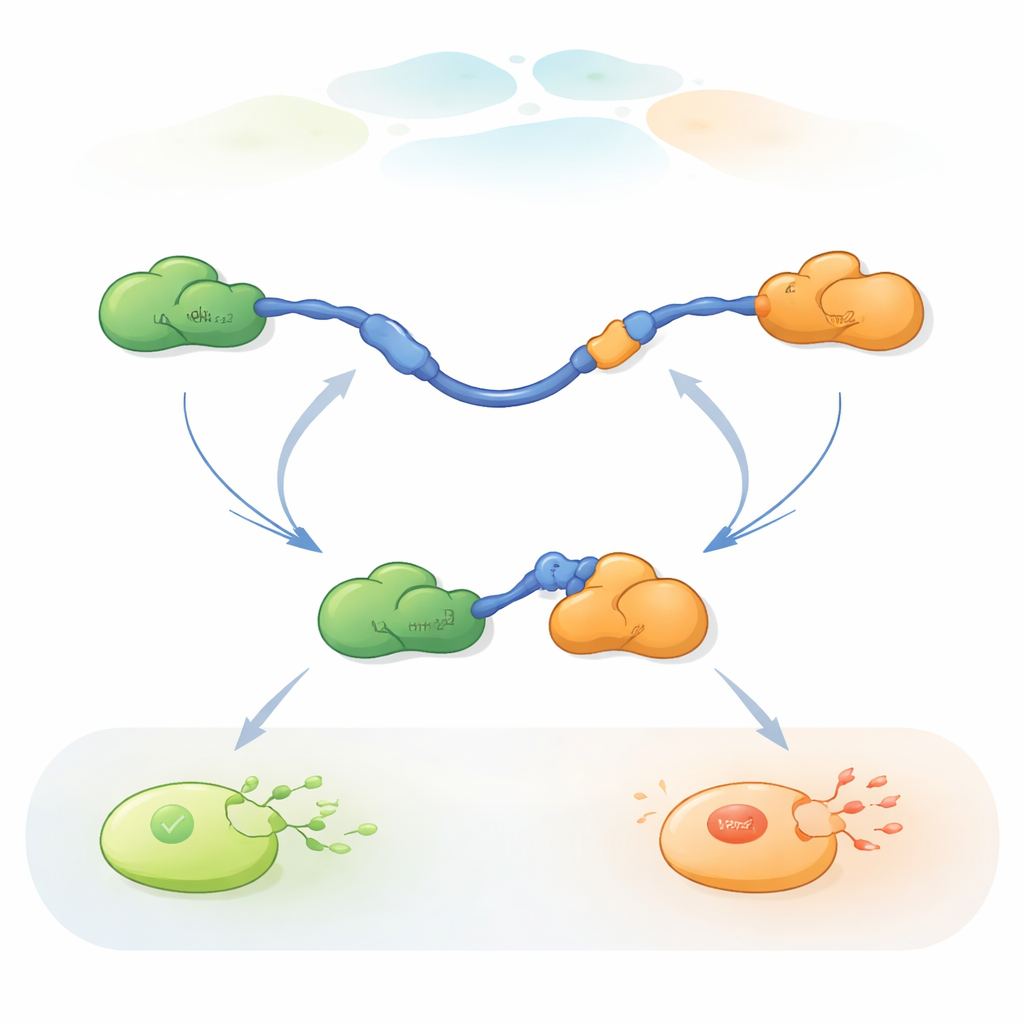
Osservare molecole singole scambiare partner
Per seguire questi rapidi scambi di partner, i ricercatori hanno utilizzato il FRET a tre colori su singola molecola, un metodo di fluorescenza che misura minuscoli cambiamenti di distanza all’interno e tra proteine. Hanno attaccato coloranti di colori diversi alle estremità del TAD di p53 e a Taz2, lasciando Mdm2 non marcato. Immobilizzando il TAD su una superficie di vetro e aggiungendo i partner in quantità controllate, hanno potuto osservare singole molecole di p53 mentre legavano, rilasciavano e rilegavano Taz2 e Mdm2 nel tempo. I cambiamenti nei pattern di fluorescenza hanno rivelato se il TAD era libero, legato solo a Taz2, legato solo a Mdm2 o brevemente legato a entrambi contemporaneamente in un complesso ternario a tre proteine.
Due modi per scambiare partner
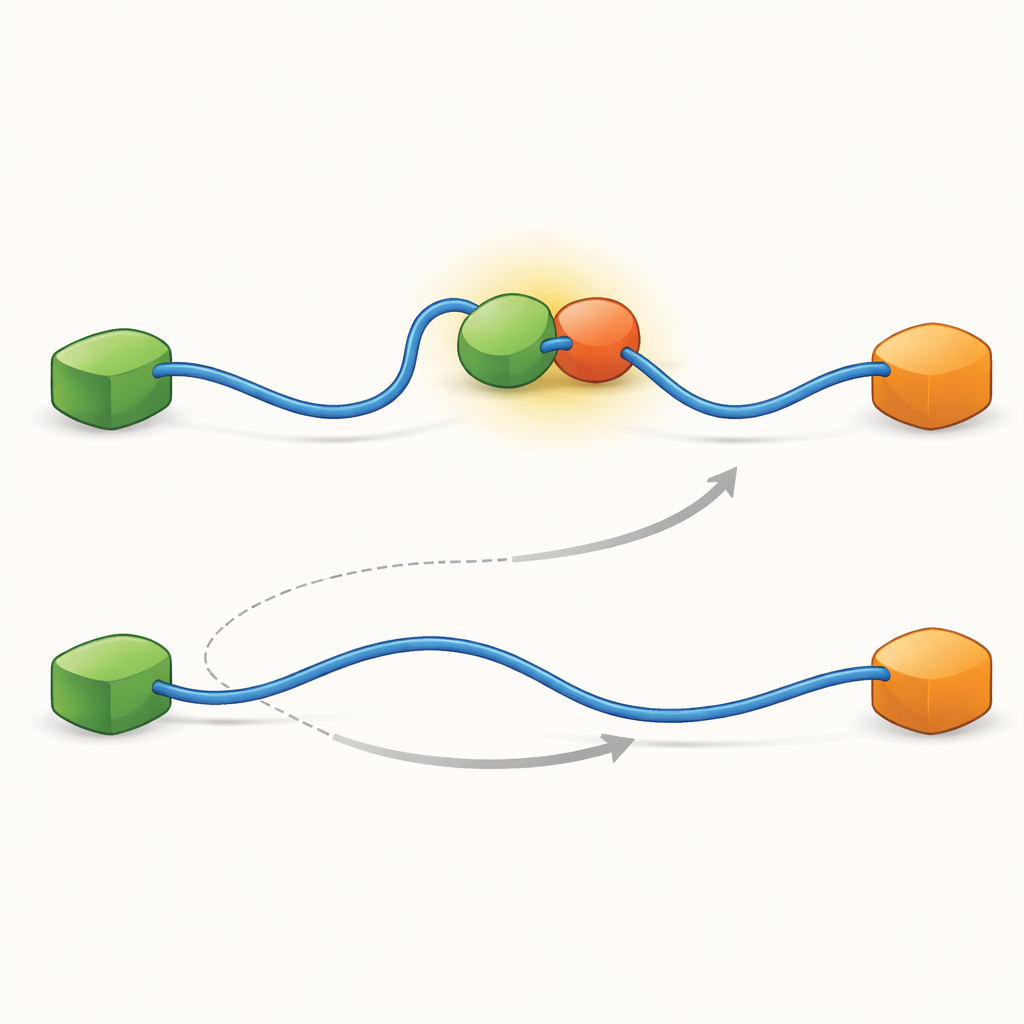
Le registrazioni hanno mostrato che il TAD può scambiare partner in due modi qualitativamente diversi. Nel percorso competitivo, il TAD si libera completamente di un partner, diventando libero e flessibile, e soltanto allora si lega all’altro partner. Nel percorso allosterico, il TAD non si stacca completamente. Piuttosto, forma un complesso ternario di breve durata in cui Taz2 e Mdm2 sono entrambi legati allo stesso tempo, ognuno aggrappandosi a una parte diversa del TAD. Da questo stato congiunto, un partner se ne va e l’altro rimane. Contando quanto a lungo il TAD rimaneva in ciascuno stato e quanto spesso avvenivano le transizioni, gli autori hanno potuto ricavare costanti di velocità per ogni passaggio e quantificare quanto frequentemente ciascun percorso veniva utilizzato in diverse condizioni.
Passaggi intermedi nascosti modellano la rete
Per scoprire cosa rende possibili questi due percorsi, il gruppo ha spinto la loro tecnica a risoluzioni temporali molto elevate e ha analizzato gli arrivi di fotoni uno per uno. Si sono concentrati sull'atto del legame del TAD a Taz2 e hanno trovato che questo processo segue a sua volta due tipi distinti di «percorsi di transizione»—brevi stadi intermedi tra lo stato non legato e lo stato completamente legato. In un tipo, Taz2 trattiene saldamente AD2 mentre AD1 resta libera; nell'altro, Taz2 si impegna più fortemente con AD1 mentre AD2 è più lasco. L'intermedio con AD2 legato permette naturalmente a Mdm2 di attaccarsi a AD1 e creare il complesso ternario, alimentando il percorso allosterico. Al contrario, l'intermedio con AD1 legato blocca Mdm2, costringendo il sistema a usare la via competitiva in cui il TAD deve staccarsi completamente prima che Mdm2 possa legarsi. Così, le forme microscopiche e fugaci che il TAD adotta durante il legame finiscono per dettare quale percorso di scambio su larga scala predomina.
Commutazione rapida per scelte di vita o di morte
Mettendo insieme tutti i numeri, gli autori mostrano che il percorso allosterico tramite il complesso ternario può portare la maggior parte del traffico tra gli stati legati a Mdm2 e quelli legati a Taz2, specialmente quando Mdm2 è abbondante, come avviene nelle cellule. Questa via permette a p53 di passare rapidamente dall’essere tenuto sotto controllo da Mdm2 all’essere attivato da Taz2 quando le condizioni cambiano, limitando al contempo il rilegarsi eccessivo di Mdm2. Modificazioni chimiche del TAD che indeboliscono il legame con Mdm2 possono spostare l’equilibrio verso il percorso competitivo, offrendo un altro livello di controllo. In termini semplici, lo studio rivela che un segmento proteico flessibile sfrutta la sua natura camaleontica per costruire una piccola stretta di mano ternaria temporanea che accelera lo scambio di partner. Questo meccanismo potrebbe essere un modo generale con cui le regioni proteiche disordinate gestiscono reti di segnalazione complesse e aiutano le cellule a rispondere rapidamente allo stress.
Citazione: Kim, JY., Chung, H.S. Dynamic control of IDP interaction network via diverse binding pathways. Nat Commun 17, 3249 (2026). https://doi.org/10.1038/s41467-026-69946-4
Parole chiave: proteine intrinsecamente disordinate, regolazione di p53, reti di legame proteico, commutazione allosterica, FRET a singola molecola