Clear Sky Science · ja
膜上スーパーコンプレックスにおける電子輸送のための表面拡散経路を導く一過性タンパク質構造
細胞はどうやって“電力線”を安全に保つか
あなたの体の細胞は毎秒数十億もの電子を移動させてエネルギー分子ATPを作り出しています。その電子の一部でも漏れると、反応性の高い化学種が生じてDNA、タンパク質、脂質を損傷します。本研究は、ミトコンドリア内膜上の小さな“電力線”タンパク質が、電子を一つのステーションから次へ安全かつ効率的に導く仕組みを掘り下げ、柔軟で一見無秩序に見えるタンパク質領域と特殊な膜脂質の意外な役割を明らかにします。
ミトコンドリア内部の忙しい電力網
ミトコンドリアの内部では、エネルギー生産は膜に埋め込まれた大型タンパク質複合体の連鎖によって進みます。重要な2つのステーション、複合体IIIと複合体IVは、小さな搬送体タンパク質シトクロムcを介して電子を受け渡します。従来はシトクロムcが水性の内部を三次元的に漂い、それが各複合体に衝突して受け渡しを行うと考えられてきました。しかし多くの生物ではこれらの複合体がスーパーコンプレックスと呼ばれるクラスターに配置されており、こうした集積が電子シャトルの効率を本当に高めるのか、それとも単に機構を整理して偶発的な漏れを減らすだけなのかが問題となっていました。
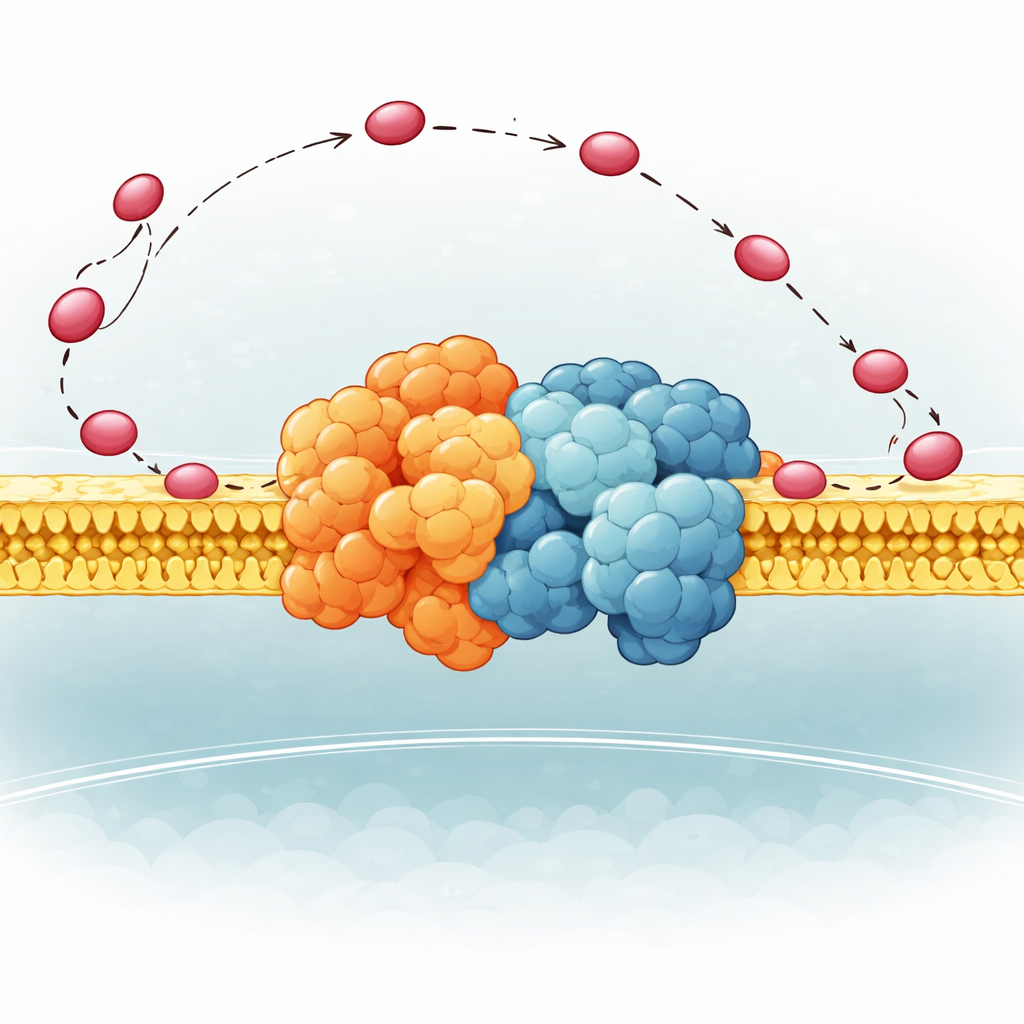
搬送体を制御する柔軟なヒンジ
筆者らは酵母の複合体IIIにある小さな部分質量QCR6に注目しました。QCR6は長く負に帯電した“ヒンジ”セグメントを持ち、その構造はこれまで柔軟すぎて明確に捉えられていませんでした。タンパク質折り畳みアルゴリズム、分子動力学、クライオ電子顕微鏡を組み合わせた高度な手法により、膜中で完全な複合体III–複合体IVスーパーコンプレックスに結合した際にこのヒンジが取りうる現実的な形状のアンサンブルを生成しました。彼らはQCR6が静止していないことを見出しました:シトクロムcが近づくとヒンジの一部が一時的にヘリックス構造を取り、搬送体を捕らえます;シトクロムcが複合体IIIから複合体IVへ移動するにつれてヒンジは伸びて部分的にほどけ、搬送体を膜表面に沿って追従し導くのです。これによりシトクロムcが周囲の流体中へ自由に漂うのではなく、表面に沿って移動するようになります。
見えない案内役としての膜脂質
膜自体も能動的な役割を果たします。研究チームはモデル内の一般的な脂質を、健康なミトコンドリアで重要とされ、心疾患や神経変性でしばしば変化する負に帯電した脂質カルジオリピンに置き換えました。シミュレーションはQCR6の正に帯電したスポットがカルジオリピンのヘッドグループに結合し、ヒンジを膜に係留することでシトクロムcをエスコートしていることを示しました。この結合により、搬送体が三次元的にランダムに飛び移るよりも、複合体IIIから複合体IVへ表面に沿って滑る方がエネルギー的に有利になります。カルジオリピンを除くと、この移動のエネルギーコストは急増し、より多くのシトクロムcがバルク溶液へ漂い、電子受容部位へ偏った配置が失われました。
ランダムな漂流から導かれるサーフィングへ
大規模なブラウン運動シミュレーションにより、研究者たちはシトクロムcがスーパーコンプレックス周辺にどこで滞留しやすいかを追跡しました。QCR6ヒンジとアニオン性脂質が揃っていると、多くのシトクロムc分子は膜近傍にとどまり、複合体IIIと複合体IVの生産的な接触部位の近くでより多くの時間を過ごしました。ヒンジが伸びると、シトクロムcの優勢な位置分布はドナー側からアクセプター側へと移動し、実質的に複合体表面に沿って“サーフィング”します。この誘導された表面拡散は、シトクロムcを機械の永久的な一部にしてしまうことなく、電子の受け渡しを迅速化します;搬送体は周囲のプールと自由に交換できます。
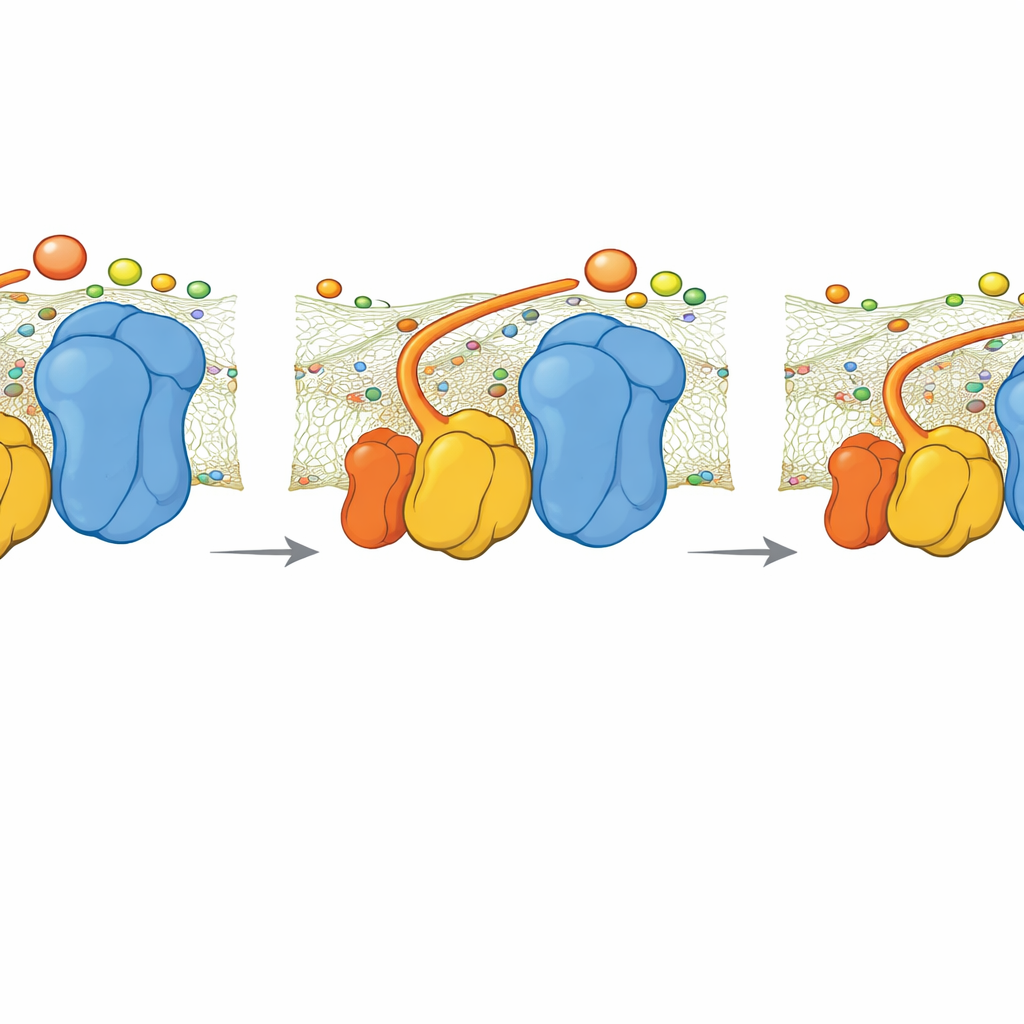
ヒンジが欠けると何が起こるか
チームはQCR6を欠く変異酵母株も調べました。クライオEM像はスーパーコンプレックスがよりコンパクトでわずかに異なる形状に再配列し、表面にわたる負電荷の連続性を部分的に回復することを示しました。それでもシミュレーションでは、QCR6ヒンジがないとシトクロムcが複合体IIIから複合体IVへ電子を移す速度が低下することが明らかになりました。電子流をATP産生に結びつける単純な動力学モデルは、高いエネルギー需要下でこの誘導喪失がATP生産を概ね30%程度削る可能性があることを示唆します。需要が低い場合は差は小さく、遅くて秩序だっていない拡散でも十分であるためです。
健康と進化にとっての意義
総じて、この研究は新たな像を支持します:柔軟なタンパク質セグメントと帯電脂質は静的な足場ではなく協調して電子搬送体を膜表面に閉じ込め、その動きを安全で生産的な経路へ偏らせている、というものです。酵母では異常に長いQCR6ヒンジと豊富なカルジオリピンがこの“再折り畳み誘導拡散”を生み出し、電子漏れを抑えエネルギー産出を高めます。他の生物では、短いヒンジや異なるスーパーコンプレックス設計、あるいは共有結合的な係留が同様の目的を果たしているように見えます。これらの微妙な誘導メカニズムを理解することは、カルジオリピンやスーパーコンプレックスの異常が加齢、心不全、神経変性で現れる理由を説明する助けになり、最終的には細胞のエネルギー代謝を調整する戦略の開発につながる可能性があります。
引用: Chan, C.K., Nguyen, J., Hryc, C.F. et al. Transient protein structure guides surface diffusion pathways for electron transport in membrane supercomplexes. Nat Commun 17, 2892 (2026). https://doi.org/10.1038/s41467-025-67110-y
キーワード: ミトコンドリア超複合体, シトクロムcの拡散, カルジオリピン, 電子伝達系, QCR6 ヒンジ