Clear Sky Science · he
מבנים של הטרטמרים חלקית מאוכלסים מספקים תובנות על הפעלה והתישנות של קולטן קאינת
למה מתגי מוח זעירים חשובות
קולטני קאינת הם מתגי חלבון מזעריים שעוזרים לתאי המוח לתקשר זה עם זה. הם מגיבים למוליך העצבי גלוטמט ומעורבים בתהליכים שמטווחים מלמידה וזיכרון ועד כאב ודלקת. כשמתגים אלה מתקלקלים הם נקשרו להפרעות כמו אפילפסיה ומחלות פסיכיאטריות. מחקר זה בוחן, בפרטי מבנה חסרי תקדים, כיצד סוג מרכזי אחד של קולטני קאינת נדלק ואז "מכבה את עצמו" כאשר רק חלק מהתת-יחידות שלו קשורות למולקולות בדמויית גלוטמט — מצב שסביר שהוא משקף מה שקורה במקומות הסינפטיים הרבים במוח.
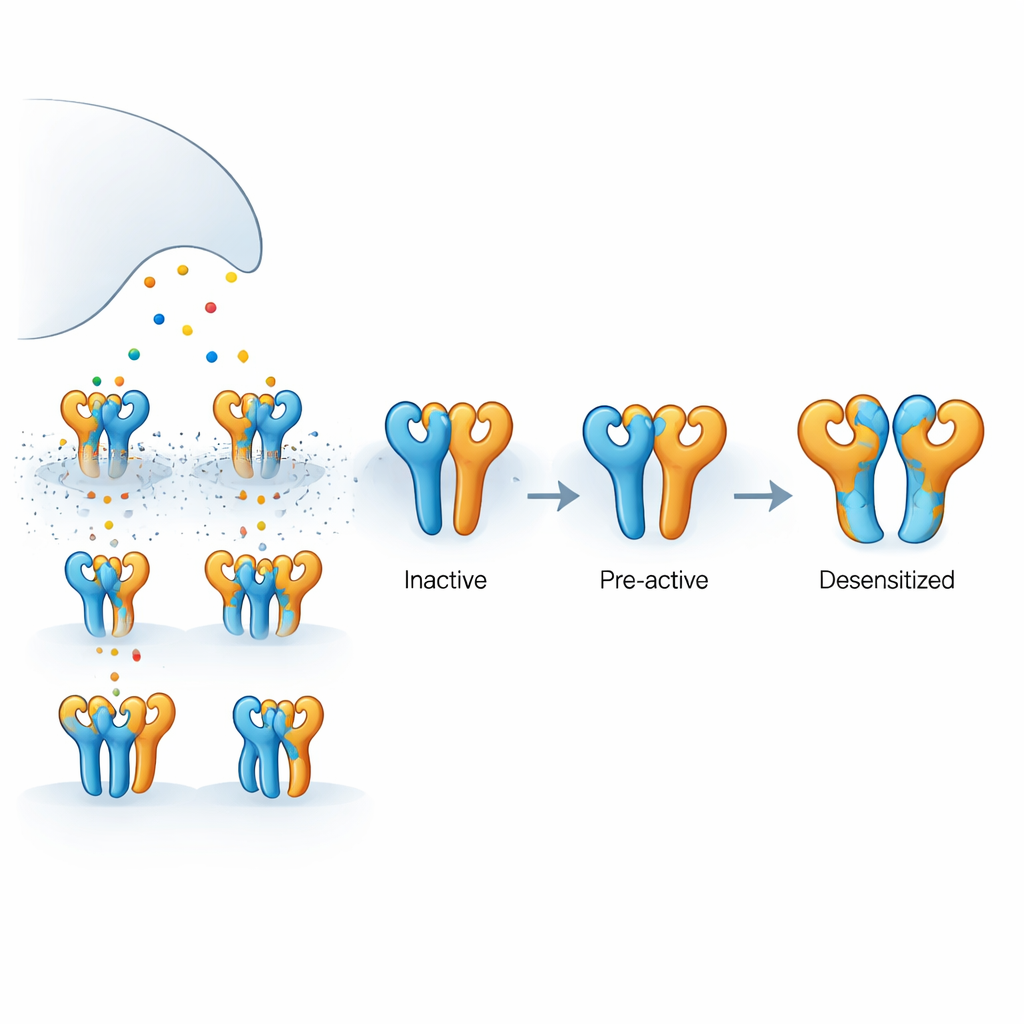
איך הקולטן הזה בנוי
הקולטן שנבדק כאן הוא קולטן קאינת המורכב משני סוגי תת-יחידות חלבוניות, הנקראות GluK2 ו-GluK5, המורכבות למבנה ארבע-חלקי. GluK5 קושרת גלוטמט בחוזקה יחודית ונוטה לתפוס שתי עמדות ספציפיות בטרטמר, בעוד GluK2 תופסת את השניים האחרים. לכל תת-יחידה יש תחום חיצוני המלכד את הגלוטמט ואזור פנימי היוצר נקב שמאפשר ליינות לזרום וליצור אותות חשמליים. מכיוון שהגלוטמט משתחרר בקצרה ונלקח במהירות, לא כל ארבע התת-יחידות תמיד מאוכלסות בעת ובעונה אחת. הבנת מה קורה כאשר רק חלק מהן קשורות היא המפתח להסברת האופן שבו סינפסות במציאות מווסתות את עוצמת האות ואת זמנו.
טריגרים שנלחצים חלקית שאינם תקועים
באמצעות רישומים חשמליים בתאים אנושיים הראו החוקרים כי תרופה סלקטיבית ל-GluK5, 5-יודו-ווילרדין, מגרמת לזרם יציב דרך קולטני GluK2/GluK5 מבלי לגרום להם "להתישן" — כיבוי מגן עצמי שלרוב בא בעקבות ההפעלה. לעומת זאת, המוליך הטבעי גלוטמט גורם לאותם קולטנים להידלק ואז להשתתק במהירות רבה. תרופה נוספת, AMPA, גם מפעילה קולטנים אלה אך מייצרת רק ההתישנות חלקית. התנהגויות אלה רומזות שכאשר רק תת-יחידות GluK5 מאוכלסות, הקולטן יכול להיפתח או להתקרב לפתיחה מבלי להיכנס למצב לא פעיל ארוך-טווח, בעוד שקישור רחב יותר דוחף את הקולטן לתצורת כיבוי עמוקה יותר.
דקות של חלקים בתנועה
כדי לראות כיצד מצבים פונקציונליים שונים אלה נראים בתלת-ממד השתמשו החוקרים במיקרוסקופיה אלקטרונית קפואה (cryo-EM) כדי ללכוד תמונות מבניות תחת תנאי ליגנד מבוקרים בקפידה. בנוכחות האגוניסט הסלקטיבי ל-GluK5 הם מצאו שתי קונפורמציות עיקריות. באחת מהן ארבעת תחומי קשירת הליגנד יצרו זוגות שלמים, עם יחידות GluK5 סגורות סביב הליגנד ויחידות GluK2 שנשארו פתוחות. מצב "שלם" זה הראה רופפות וסיבוב עדינים בין התחומים המזווגים והתרחבות מדידה של האלפאות היוצרות את הנקב הפנימי, בהתאם לקונפיגורציה מוקדמת-פעילה, מוכנה להיפתח אף על פי שהתעלה היונית עדיין הייתה סגורה. במצב השני, "חלקית קרועה", זוג אחד של תחומים נפרד בעוד השני נשאר שלם, מה שמרמז על ביניים בדרך אל ההתישנות המלאה אך שאינו נעול עדיין.
צירי סתר שמכוונים את התזמון
על ידי השוואת מבנים אלה עם קולטני קאינת שנפתרו בעבר במצב פעיל מלא ומושתק מלא, המעקב אחר תנועות במעטפת החיצונית אל התעלה נעשה אפשרי. הם זיהו אזורים מרכזיים של מגע הן בתוך זוגות התחומים והן בין הזוגות שפועלים כצירים וחיזוקים, במיוחד סביב שתי תת-יחידות GluK5 היושבות זו מול זו במבנה. שינויי מיקום עדינים באתרים אלה שינו עד כמה התחומים מחזיקים זה את זה וכיצד הם מושכים את הקישורים הגורמים לפתיחת הנקב. כאשר החוקרים החליפו חומצות אמינו מסוימות בממשקים אלה הם הצליחו לגרום לקולטן להשתתק מהר יותר או לאט יותר ולשנות עד כמה הוא נוטה להתישנות. זה מסביר מדוע קולטנים המכילים GluK5, במיוחד, נשארים פעילים לזמן בלתי רגיל לאחר התפרצות קצרה של גלוטמט.
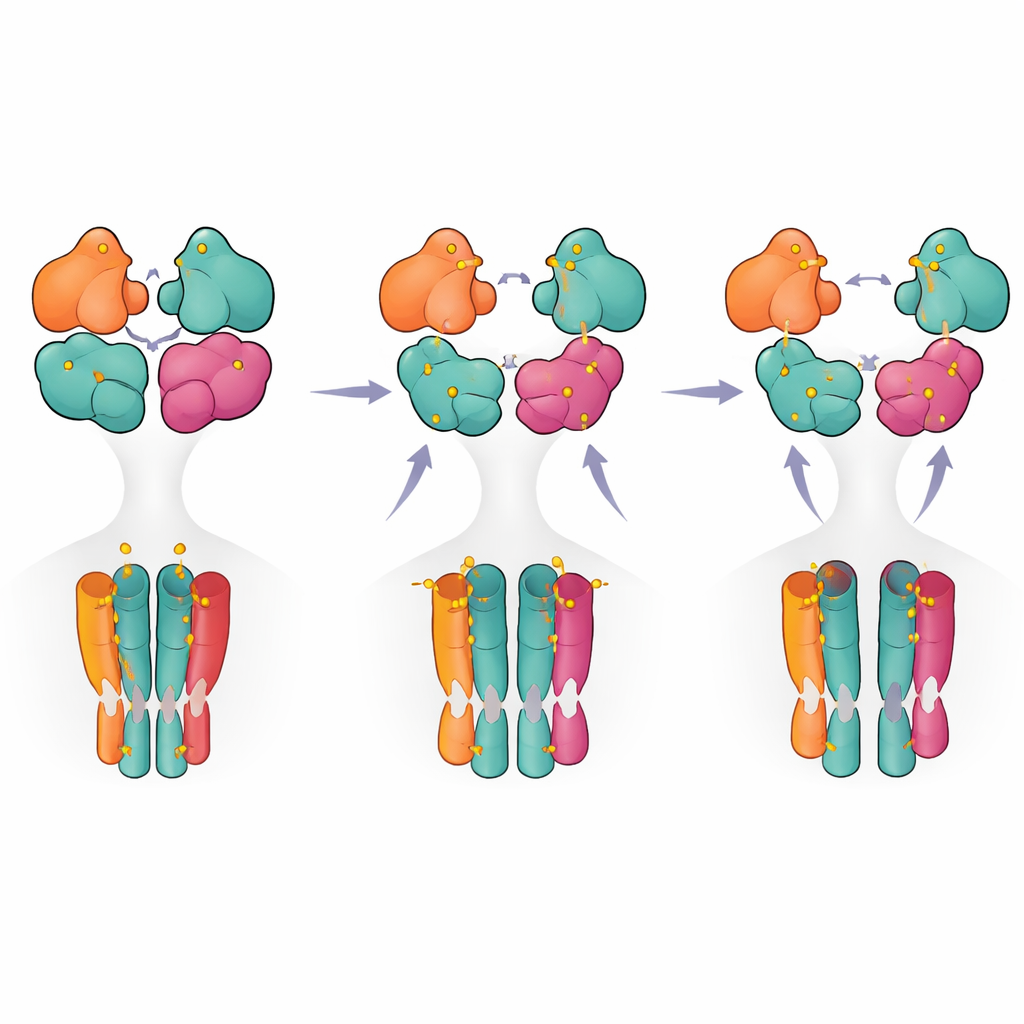
מפת דרכים צעד-אחר-צעד מהפעלה לכיבוי
בשילוב התמונות המבניות והמדידות הפונקציונליות מציעים החוקרים מודל צעד-אחר-צעד שבו הקולטן נע ממצב מנוחה למצב טרום-פעיל כאשר רק שתי תת-יחידות (בדרך כלל זוג ה-GluK5) מאוכלסות. קשירת ליגנד נוספת מעודדת קריעה חלקית של אחד מזוגות התחומים, ורק כאשר שלוש או ארבע תת-יחידות מאוכלסות מגיע הקולטן לתצורה קרועה לחלוטין המקושרת להתישנות ממושכת. עבור קוראי-לא-מומחים משמעות הדבר היא שהקולטן מתנהג פחות כמו מתג פשטני על-מול-ל-off ותפא יותר כמו דימר רב-שלבי, עם דפוסי תפוסה ומגעים פנימיים שונים המכוונים את עוצמת וזמן האיתות. תובנות אלה לא רק מאירות את הפעולה הבסיסית של קולטן מוח מרכזי אלא גם מדגישות "נקודות חמות" מבניות שעשויות להיות יעדים לתרופות עתידיות שמטרתן לכוונן באופן עדין את פעילות קולטני הקאינת במחלות.
ציטוט: Khanra, N.K., Strauss, A., Moreno Wasielewski, L. et al. Structures of partially occupied hetero-tetramers provide insight into kainate receptor activation and desensitization. Nat Commun 17, 3789 (2026). https://doi.org/10.1038/s41467-026-72226-w
מילות מפתח: קולטני קאינת, אותות גלוטמט, שידור סינפטי, מבני cryo-EM, התישנות קולטן