Clear Sky Science · he
קיפול קו-שיעתוק מארגן חישה סדרתית מרובת-אפקטורים על ידי ריבוסוויץ׳ טנדם לגליצין
איך מתגי RNA עוזרים לחיידקים לקבל החלטות חכמות
חיידקים חיים בסביבות שמשתנות בהתמדה, ובכל זאת צריכים להחליט בזמן אמת אילו גנים להפעיל או לכבות כדי לחסוך אנרגיה ולהישרדות. המחקר הזה מגלה כיצד "מתג" RNA זעיר בתוך החיידק Bacillus subtilis מאזין למספר איתותים מולקולריים בו-זמנית — חומצת אמינו, יוני מתכת וחלבון-קו-פקטור — ואז בוחר במהירות האם להמשיך בקריאת הגן או לעצור אותה. הבנת מעגל ההחלטה המוכל זה לא רק חושפת חוקים בסיסיים של בקרה גנטית, אלא גם עשויה להשריש כלים חדשים לביולוגיה סינתטית ולעיצוב אנטיביוטיקות.
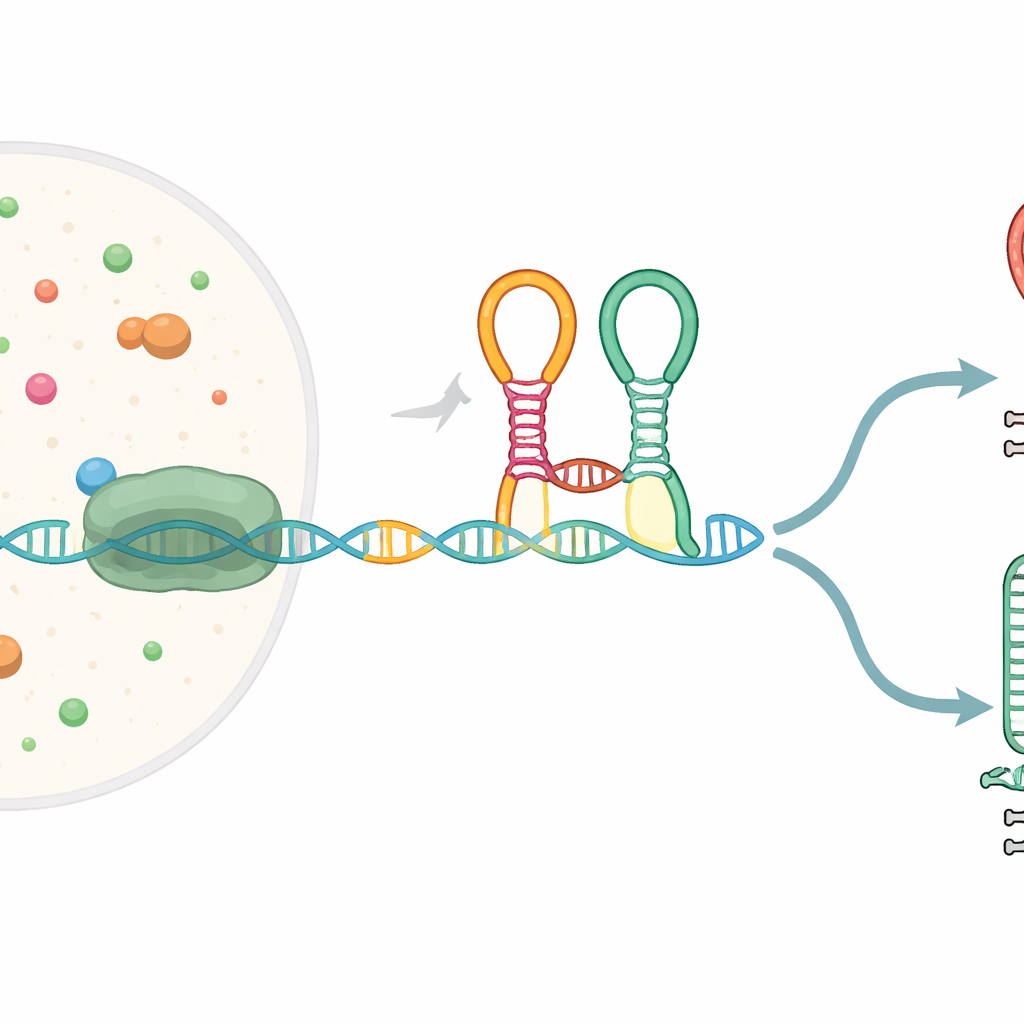
חיישן זעיר המשולב בהודעת הגן
רבים מהגנים החיידקיים מתחילים בקטעי RNA מיוחדים הנקראים ריבוסוויצ׳ים, שמתקפלים לצורות שיכולות לחוש מולקולות קטנות ובתגובה לשנות ביטוי גנים. הריבוסוויץ׳ הנחקר כאן, הידוע כטנדם ריבוסוויץ׳ לגליצין, יושב מול גנים המסייעים בפירוק חומצת האמינו גליצין. הוא מכיל שני יחידות חישה סמוכות ("אפטמרים") ששתיהן מזהות גליצין ומחוברות בעיקול חד שנקרא kink-turn. בזמן שההודעה של ה-RNA מועתקת מה-DNA על ידי RNA פולימראז, ריבוסוויץ׳ זה מתקפל תוך כדי תנועה ויכול לכוון את התהליך либо להשלמת הטרנסקריפט המלא, או לקיצורו בתחנת עצירה מובנית.
עצירות שיוצרות נקודות בדיקה להחלטה
המחברים גילו שהאנזים המעתיק RNA אינו נע חלק דרך אזור זה; במקום זאת הוא עוצר בשלוש נקודות ספציפיות בתוך הריבוסוויץ׳. העצירות האלה פועלות כמו רמזורים, ומעניקות ל-RNA המתהווה זמן להתקפל ולהתקשר עם מולקולות בסביבתו. בעצירה הראשונה נוצר האפטמר העל-זרימה ומוכן לקשור גליצין. בעצירות מאוחרות יותר צצים חלקים מהאפטמר השני וההליקס ההכרעתי הסופי. חלבון מסייע בשם NusA מאריך שתי מהעצירות הללו, במיוחד כשגליצין נוכח, ובכך מרחיב את חלון הזמן שבו הריבוסוויץ׳ יכול לחוש את סביבתו ולמחויב להמשך השיעתוק או להפסקתו.
חישה של יונים, גליצין ושותף חלבוני שלב אחרי שלב
באמצעות מיקרוסקופיית פלואורסנציה למולקולה בודדת ופרובינג כימי ברוחב-פס רחב, הצוות הראה שהריבוסוויץ׳ אינו פועל בבת-אחת. בתחילה, יוני אשלגן מייצבים את ה-kink-turn בבסיס ה-RNA ומכוונים את האפטמר הראשון לעבר נתיב הטרנסקריפט המתהווה. לאחר מכן גליצין נקשר לאפטמר הראשון ומקבע את המבנה המקומי שלו. אירועים אלה מעודדים מגעים טווח-ארוך בין שני האפטמרים, שמארגנים מראש את שתי כיסי הקישור לגליצין גם לפני שהגליצין השני מגיע. בעצירה הסופית בקרבת שער-הסיום המובנה, גליצין שני יכול להיקשר לאפטמר הש downstream בסביבה הממוינת מראש זו, ועיכוב המוגבר על ידי NusA תורם להעדפת השלמת הקונפיגורציה הקומפקטית והפעילה.
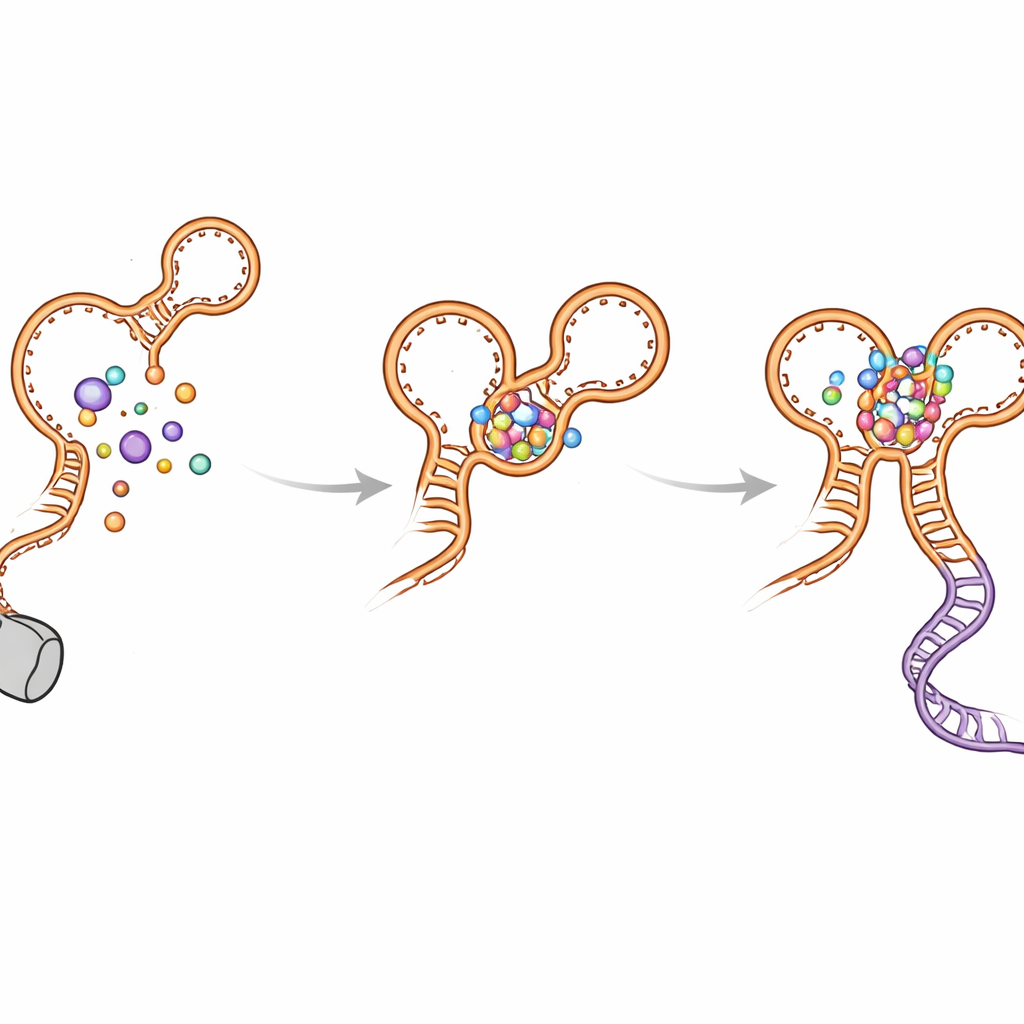
סטיות קיפול וכיצד המתג מונע הסתבכות
המחברים גילו גם מספר לולאות "סיבוב שגוי" שנוצרות זמנית כאשר ה-RNA גדל. מבנים לא-מקומיים אלה מופיעים בשני האפטמרים ועלולים לעכב את היווצרות הצורה הסופית והפעילה. ההליקס של ה-kink-turn (שנקרא P0) והמגעים בין האפטמרים עוזרים לפרק את הביניים האלו ולכוון את הקיפול לפריסה פונקציונלית. כאשר P0 או נקודות מפתח למגע מופרעות על ידי מוטציות, הריבוסוויץ׳ מאבד חלק ניכר מהרגישות והיעילות שלו: גליצין עדיין נקשר, אך העגינה לטווח-ארוך מוחלשת, ההליקס ההכרעתי פחות יציב, וקריאת השיעתוק המלאה פוחתת. סימולציות מחשב תמכו בממצאים אלה והראו שהריבוסוויץ׳ הרבה יותר קומפקטי ויציב כאשר P0 ושתי מאגרי הגליצין שלמים.
שער לוגי שאינו בשיווי משקל בתאים חיים
במבט כולל, התוצאות תומכות בתמונה שבה הטנדם ריבוסוויץ׳ לגליצין מתנהג כשער לוגי קיניטי ולא כחיישן מבוסס-שיווי משקל פשוט. במקום ששתי מולקולות הגליצין יקשרו בו-זמנית למבנה שנוצר מראש, הקישור והקיפול מתרחשים בסדר מדויק מ-5׳ אל 3׳, מנוהלים על ידי עצירות השיעתוק ומעוצבים על ידי יוני אשלגן וחלבון NusA. התהליך הסדרתי והלא-במערך הזה מאפשר לחיידק לאינטגרייט מספר אותות — מאזן יונים תאיים, רמות גליצין ומהירות השיעתוק — לפני קבלת ההחלטה האם לבטא במלואן את הגנים המפרקים גליצין שמאחורי הריבוסוויץ׳. העבודה מציעה שאותה שותפות קינטית מונעת על-ידי דינמיקה עשויה להיות אסטרטגיה כללית שבה מתגי RNA מתאמים מספר גורמים תאיים כדי לכוונן בעדינות את ביטוי הגנים.
ציטוט: Romero, R.A., Chauvier, A., Teh, S.S. et al. Co-transcriptional folding orchestrates sequential multi-effector sensing by a glycine tandem riboswitch. Nat Commun 17, 2779 (2026). https://doi.org/10.1038/s41467-026-69648-x
מילות מפתח: ריבוסוויץ׳, חישה של גליצין, קיפול RNA, עיכוב שיעתוק, ויסות גנים בחיידקים