Clear Sky Science · zh
H 型菌毛环化与菌毛生物发生具有宽容性,但静电扰动会削弱接合效率
为何微小的细菌“电缆”至关重要
细菌不断交换 DNA,共享诸如抗生素抗性等性状,这些性状可能将无害微生物变为难以治疗的威胁。一个主要的交换途径是接合:供体细菌使用一种细长、毛发般的表面“电缆”——菌毛,连接邻近细胞并传递质粒 DNA。本研究提出了一个表面看似微妙但意义重大的问题:菌毛表面的精确原子排列以及受体外膜中的脂质,如何共同决定 DNA 转移能否成功?
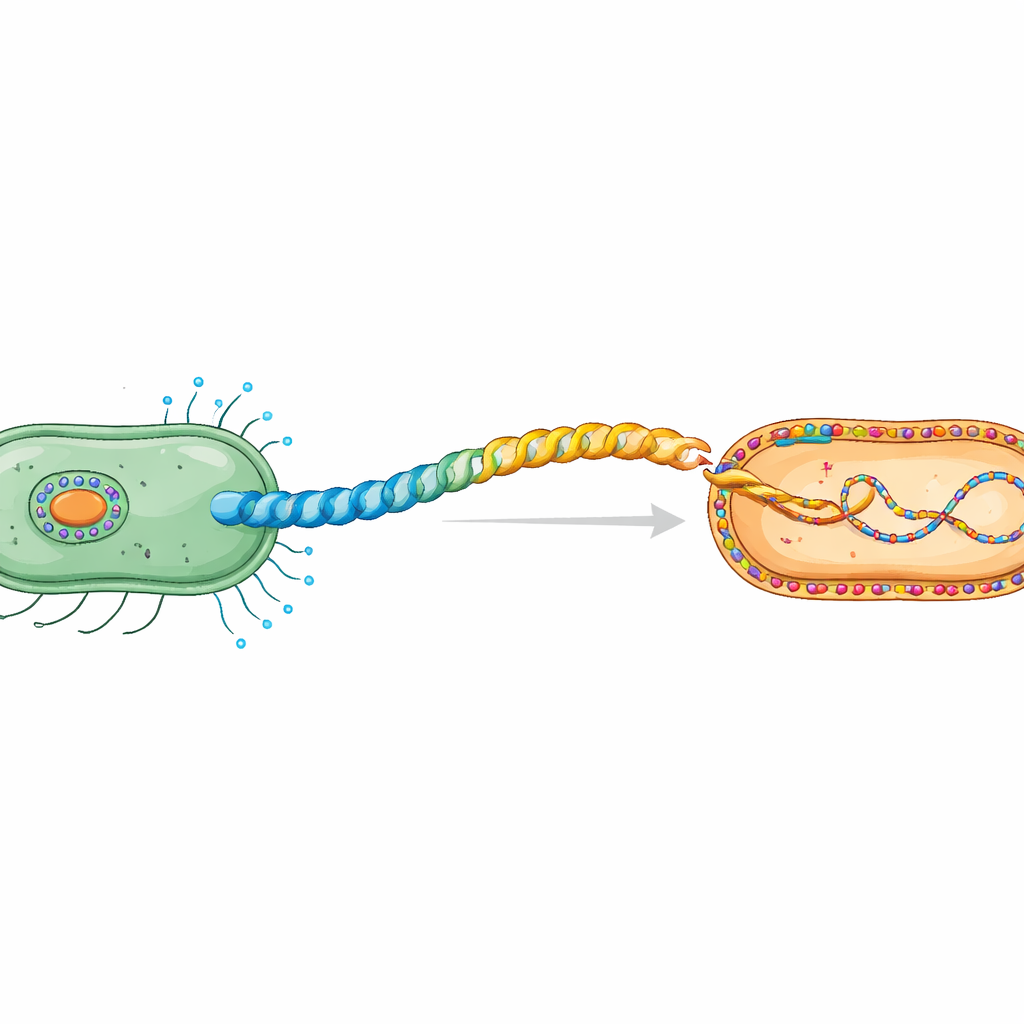
细菌细胞之间的“秘密握手”
在接合过程中,质粒 DNA 在接触依赖的机制下从供体细胞传递到受体细胞。供体细胞构建一种特殊的菌毛,这是一种由重复的菌毛蛋白单元组成的柔性螺旋管。本研究聚焦于由 IncHI1 质粒 R27 编码的 H 型菌毛,该质粒家族以传播抗生素抗性而闻名。早期冷冻电子显微镜研究显示,其菌毛蛋白 TrhA 异常:蛋白并非保持端开,而是通过将 N 端(Gly1)与 C 端(Asp69)连接起来“系成一个环”。这种环化对于形成能够转移 DNA 的功能性菌毛是必要的。
高度保守的蛋白末端,却出乎意料的化学宽容性
作者通过挖掘大型质粒数据库,分析了来自相关质粒的 147 个 TrhA 序列。尽管许多位置存在变异,但闭合环的两个残基——Gly1 和 Asp69——完全保守,这提示存在强烈的进化压力维持它们。团队随后在实验室系统性地改变了这两个位置,构建了多种突变菌毛蛋白。高分辨率冷冻电镜显示,即便将 Asp69 换成截然不同的氨基酸(如 Asn、Ala、Gly 或 Arg),TrhA 仍能完成环化并组装成在结构上几乎与野生型无异的菌毛。换言之,形成环和构建菌毛的化学步骤具有出乎意料的宽容性。
当菌毛表面电荷阻断连接时
尽管外观正常,突变菌毛在与一系列革兰氏阴性受体进行接合测试时表现出截然不同的行为。当 Asp69 被中性氨基酸(Asn、Ala、Gly)替换时,质粒仍能较好地转入大肠杆菌和克雷伯氏肺炎菌等物种。在某些其他物种中,出现了微妙的趋势:较大的侧链通常支持略高的转移效率。情况在将 Asp69 或 Gly1 替换为带正电的氨基酸(如精氨酸或赖氨酸)时完全不同。这些改变将菌毛外表从带负电翻转为强正电。尽管突变菌毛仍可形成,但与野生型菌毛相比,对常规受体的接合能力下降了多达百万倍。
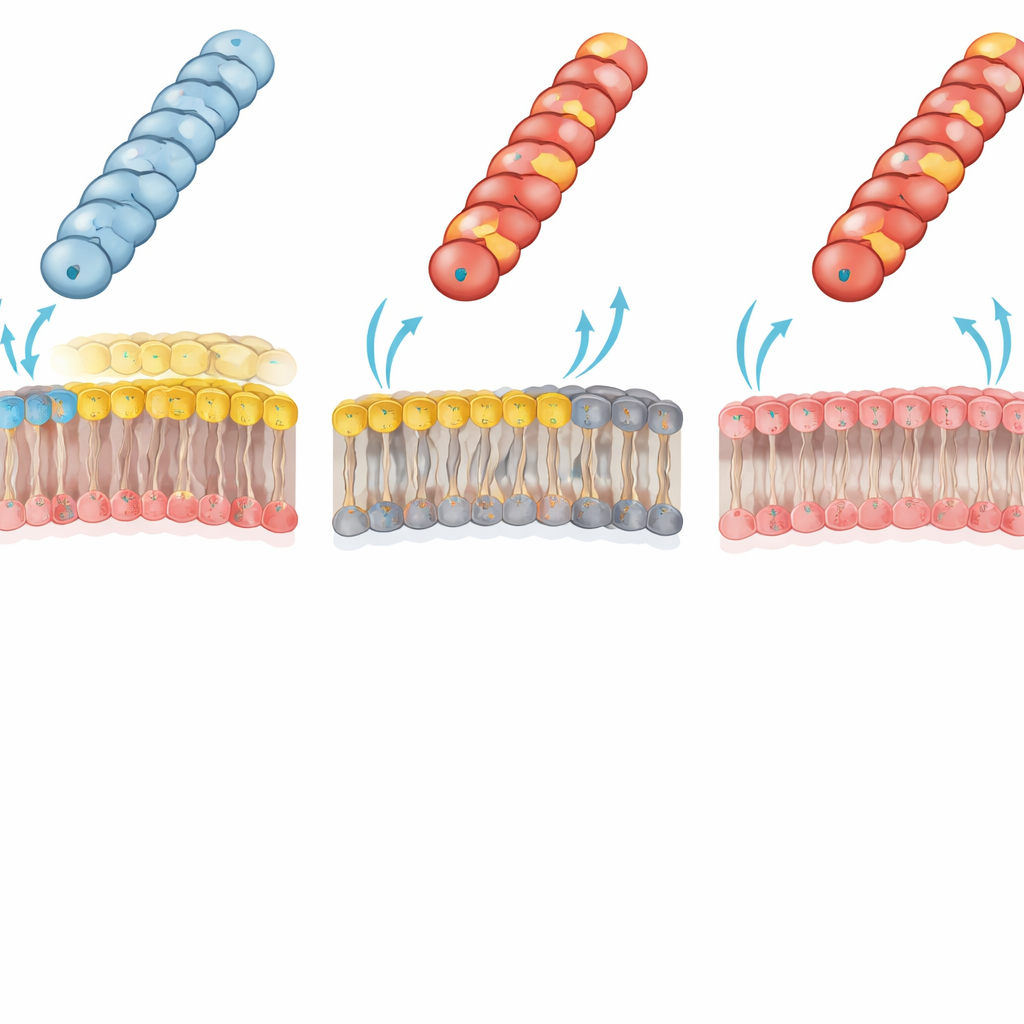
膜脂质决定谁能接收 DNA
作者将这种显著的失败归因于菌毛与受体外膜之间的静电冲突。在典型的革兰氏阴性细菌中,外膜内层含有一种主要的“两性离子”磷脂——磷脂酰乙醇胺(PE),其分子同时带有正负两类基团。团队使用了一种经过特殊改造、缺乏 PE 的大肠杆菌菌株;该菌株外膜的内层完全由带负电的脂质构成。在这个缺 PE 的受体中,先前功能受损的带正电菌毛(如 Gly1Lys、Asp69Lys 或 Asp69Arg)现在几乎能像野生型一样高效地转移质粒。这一逆转有力支持了这样一种观点:带正电的菌毛表面与富含 PE 的膜之间的不利电荷–电荷相互作用,通常会阻止菌毛端部采取适合高效 DNA 转移的接触构象。
静电微调如何塑造抗生素抗性传播
综合来看,结果揭示了明确的分工。将菌毛蛋白链切断并重新连接以形成环的化学机制相当灵活——许多不同的末端残基仍能被缝合并组装成外观正常的菌毛。但成功的接合对这些暴露残基的体积,尤其是电荷,极为敏感,因为它们决定了菌毛表面的电势,这一电势必须与受体膜的脂质环境匹配。这种静电“微调”有助于解释为何 Gly1 和 Asp69 在自然界中高度保守。它也暗示,膜脂成分的变化——受环境压力或宿主条件驱动——可能会调节细菌交换携带抗生素抗性基因质粒的容易程度。实际上,细菌“握手”的双方会共同进化它们的表面,以平衡基因共享的利益与风险。
引用: He, S., Ishimoto, N., Wong, J.L.C. et al. H pilin cyclisation and pilus biogenesis are promiscuous but electrostatic perturbations impair conjugation efficiency. Nat Commun 17, 2888 (2026). https://doi.org/10.1038/s41467-026-69599-3
关键词: 细菌接合, 抗生素抗性, 菌毛结构, 膜脂质, 静电相互作用