Clear Sky Science · sv
En heteromerisk TRP-kanal som fungerar som en WNT‑aktiverad G‑protein‑kopplad receptor
Varför ett njurprotein är viktigt för alla
Autosomalt dominant polycystisk njursjukdom (ADPKD) är en av de vanligaste ärftliga njursjukdomarna och en ledande orsak till njursvikt i hela världen. Under årtionden har forskare känt till de viktigaste generna som är inblandade—PKD1 och PKD2—men inte vad deras proteinprodukter egentligen gör i cellerna. Denna studie visar att dessa proteiner bildar en ovanlig receptor i cellmembranet som kommunicerar direkt med klassiska signalomkopplare inuti cellen. Genom att förklara hur detta system normalt håller en nyckelbudbärarmolekyl i schack ger arbetet en tydligare bild av hur cystor uppstår vid ADPKD och pekar på nya möjligheter för behandling.
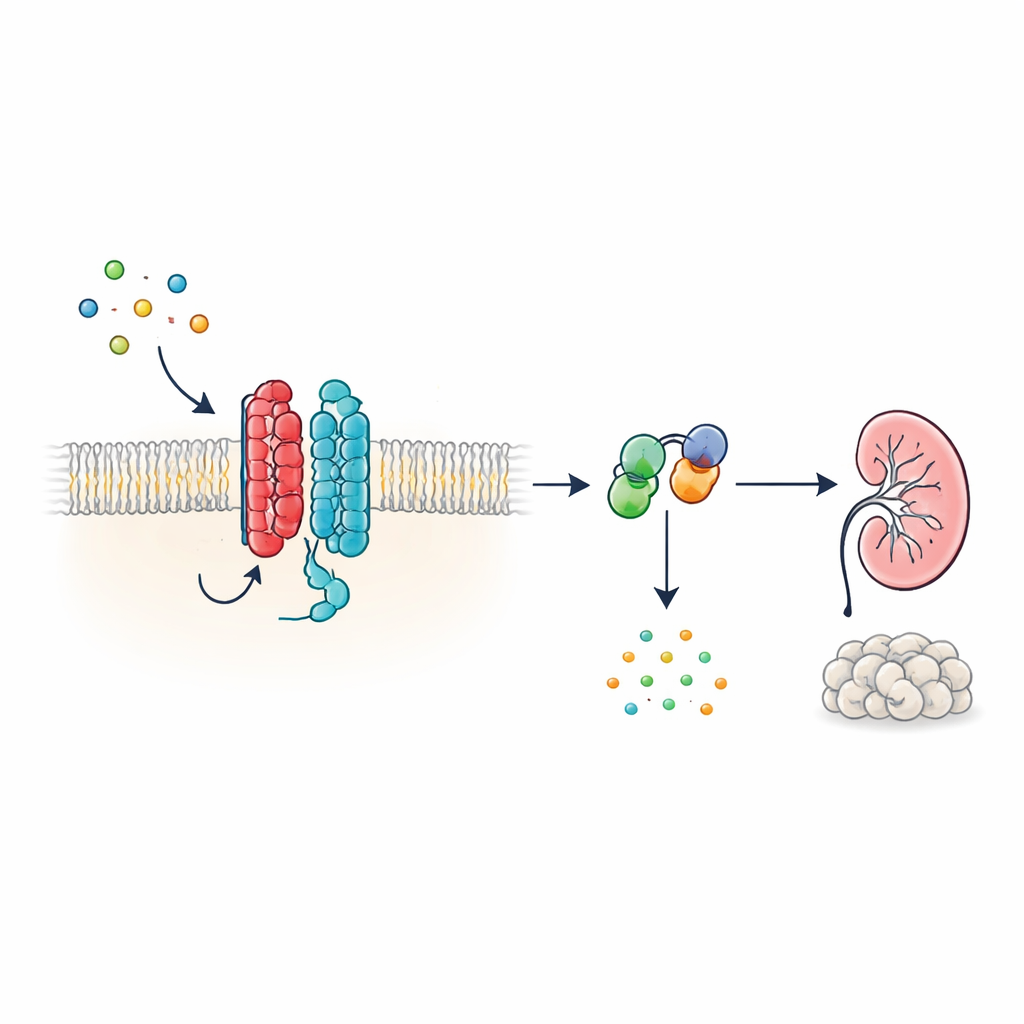
En ny typ av cellulär antenn
Många hormoner och signaler i kroppen verkar via G‑protein‑kopplade receptorer (GPCR), en mycket stor familj av ytproteiner med en karakteristisk sju‑segmentstruktur som går genom cellmembranet. PKD1 ser emellertid annorlunda ut: det har 11 segment och har klassificerats tillsammans med jonkanaler snarare än klassiska receptorer. Författarna visar att PKD1, trots denna okonventionella form, beter sig som en äkta GPCR när det paras med PKD2. Tillsammans bildar de två proteinerna ett komplex i membranet som svarar på WNT‑proteiner—mångsidiga signalmolekyler som är involverade i utveckling och vävnadsunderhåll. När WNT binder PKD1 aktiverar komplexet intracellulära G‑proteiner och förändrar nivåerna av cykliskt AMP (cAMP), en liten molekyl som starkt påverkar celltillväxt och vätskeutsöndring.
Hur WNT‑signaler färdas genom PKD1–PKD2‑komplexet
För att kartlägga denna signalväg använde forskarna ljusbaseda biosensorer som rapporterar hur G‑proteiner omorganiseras i levande celler. I humana njuravledda celler konstruerade för att uttrycka PKD1 fick flera olika WNT‑familjemedlemmar specifika G‑proteinsubenheter att splittras från sina partners—en klassisk markör för aktivering. Svaret var starkast för en inhibitorisk G‑proteinsubtyp, känd som Gαi3, och involverade även närliggande Gαi1, Gαi2 och Gαq‑subenheter, men inte de G‑proteiner som typiskt höjer cAMP. Ytterligare biosensorer visade att WNT‑stimulering inte bara förde G‑proteiner i nära kontakt med PKD1, utan också drev den kemiska steget där GDP byts mot GTP på Gα‑subenheten, vilket fullbordar aktiveringscykeln. Dessa effekter uppträdde även när andra kända WNT‑receptorer var nedtryckta, vilket tyder på att det är PKD1 självt, snarare än klassiska WNT‑receptorer, som är ansvarigt.
PKD2 som den nödvändiga partnern och grindvakten
Även om PKD1 kan binda både WNT‑ligander och G‑proteiner, verkar det inte ensamt. Gruppen använde genredigering för att ta bort PKD2 från celler och fann att WNT inte längre kunde driva G‑proteinaktivering via PKD1. Återinförande av normalt PKD2 återställde signaleringen, men sjukdomsassocierade PKD2‑varianter som stör komplexets uppbyggnad eller membranriktning gjorde det inte. Mikroskopi och energitransfermätningar visade att PKD2 fungerar som en chaperon, eskorterar PKD1 till cellytan och stabiliserar det där. Viktigt är att mutationer som ändrar PKD2:s jonkanalaktivitet utan att påverka dess partnerskap med PKD1 lämnade G‑proteinsignaleringen intakt, och att blockera jonflöde med en kemisk hämmare hade liten effekt. Detta tyder på att PKD1–PKD2‑komplexet i detta sammanhang fungerar främst som en receptor snarare än som en aktiv jonkanal.
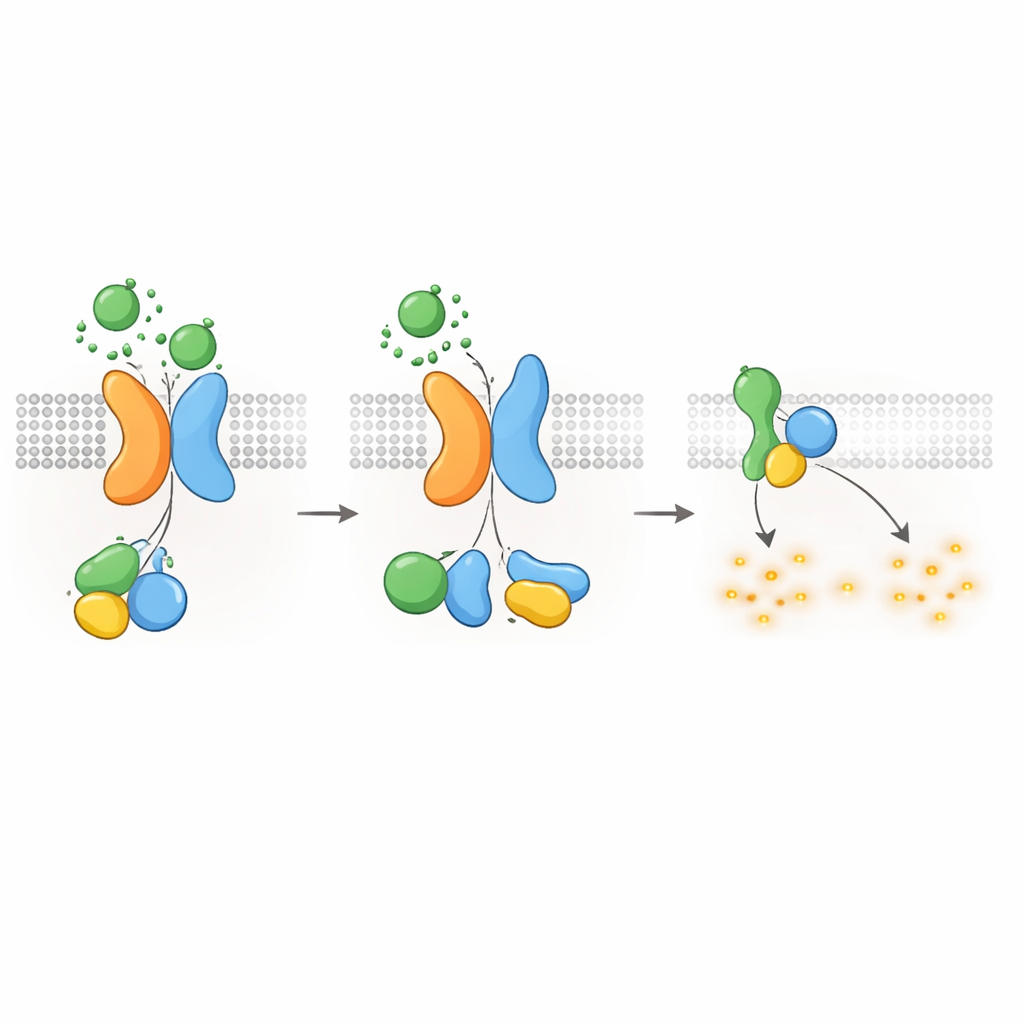
Att hålla cAMP—och cysttillväxt—under kontroll
Eftersom Gαi‑typens G‑proteiner är kända för att dämpa cAMP‑produktion frågade författarna nästa om PKD1–PKD2‑receptorn direkt kunde sänka cAMP‑nivåer. Med en luminescerande reporter som följer cAMP i realtid visade de att WNT‑stimulering i PKD1‑uttryckande celler bromsade uppstigningen av cAMP som utlösts av forskolin, en förening som ökar cAMP‑produktionen. WNT sänkte också vilande cAMP‑nivåer, och båda effekterna blockerades av pertussistoxin, vilket specifikt inaktiverar Gαi‑proteiner. Sjukdomsframkallande PKD1‑mutationer—som påverkar WNT‑bindning, autoproteolytisk klyvning eller G‑proteinsinteraktion—försvagade kraftigt denna cAMP‑sänkande respons. Celler utan PKD2 visade inte heller WNT‑beroende cAMP‑undertryckning, om inte normalt PKD2 återinfördes. Dessa fynd kopplar de molekylära felen i PKD1 eller PKD2 direkt till förlust av cAMP‑kontroll, en central drivkraft för cysttillväxt vid ADPKD.
Att stänga av signalen och flytta in receptorer
Liksom andra GPCR:er måste PKD1–PKD2‑komplexet stängas av och återvinnas efter aktivering. Författarna fann att ett kinas kallat GRK6, som finns vid plasmamembranet, associerar starkt med PKD1 när PKD2 är närvarande och hjälper till att ta bort komplexet från cellytan. Ett annat protein, β‑arrestin 2, rekryteras sedan till det aktiverade komplexet och eskorterar både PKD1 och PKD2 in i tidiga endosomer inne i cellen. Blockering av GRK6‑aktivitet eller β‑arrestin‑funktion minskade denna internalisering och återställde delvis G‑proteinsignaleringen. Dessa desensitiseringssteg tyder på att PKD1–PKD2‑receptorn är tätt reglerad, för att förhindra okontrollerad dämpning av cAMP under normala förhållanden.
Vad detta betyder för njursjukdom och vidare
Tillsammans etablerar resultaten PKD1–PKD2‑komplexet som en ny klass av G‑protein‑kopplad receptor—en okonventionell 11‑transmembranreceptor som är beroende av en partnersubenhet för korrekt placering vid cellytan. I friska njurar aktiverar WNT‑signaler via denna receptor inhibitoriska G‑proteiner och håller cAMP‑nivåerna inom ett säkert intervall, vilket hjälper till att bevara normal tubulär diameter och förhindra cystbildning. När PKD1 eller PKD2 är muterade sviktar denna WNT–GPCR‑väg, cAMP ökar och cystor expanderar. Genom att klargöra detta tidiga steg i sjukdomsprocessen öppnar studien möjligheter för terapier som syftar till att återställa eller efterlikna PKD1–PKD2‑signalering, vilket potentiellt kan erbjuda mer precisa alternativ till nuvarande läkemedel som mer brett dämpar cAMP.
Citering: Hardy, E.P., Haider, A.N., Patel, M.M. et al. A heteromeric TRP channel that functions as a WNT-activated G protein-coupled receptor. Nat Commun 17, 3233 (2026). https://doi.org/10.1038/s41467-026-69932-w
Nyckelord: polycystisk njursjukdom, PKD1 PKD2, G‑protein‑kopplad receptor, WNT‑signalering, cAMP