Clear Sky Science · he
ערוץ TRP הטרומרי שפועל כקולט זוג-חלבוני פעיל על-ידי WNT
מדוע חלבון כלייתי חשוב לכולנו
מחלת הכליה הפוליציסטית התורשתית בדומיננטיות אוטוסומלית (ADPKD) היא אחת המחלות התורשתיות השכיחות של הכליות וסיבה מרכזית לאי-ספיקת כליות ברחבי העולם. במשך עשורים ידעו המדענים מהם הגנים המרכזיים המעורבים—PKD1 ו‑PKD2—אך לא ידעו מה עושים חלבונים אלה בתוך התאים. המאמר הזה מגלה שהחלבונים יוצרים קולט יוצא דופן על פני התא שמתקשר ישירות למתגים הסיגנליים הקלאסיים בתוך התא. על ידי הסבר האופן שבו מערכת זו שומרת על מולקולת שליח מרכזית תחת שליטה, העבודה מציעה תמונה ברורה יותר של אופן היווצרות הציסטות ב‑ADPKD ומצביעה על דרכים חדשות לטיפול במחלה.
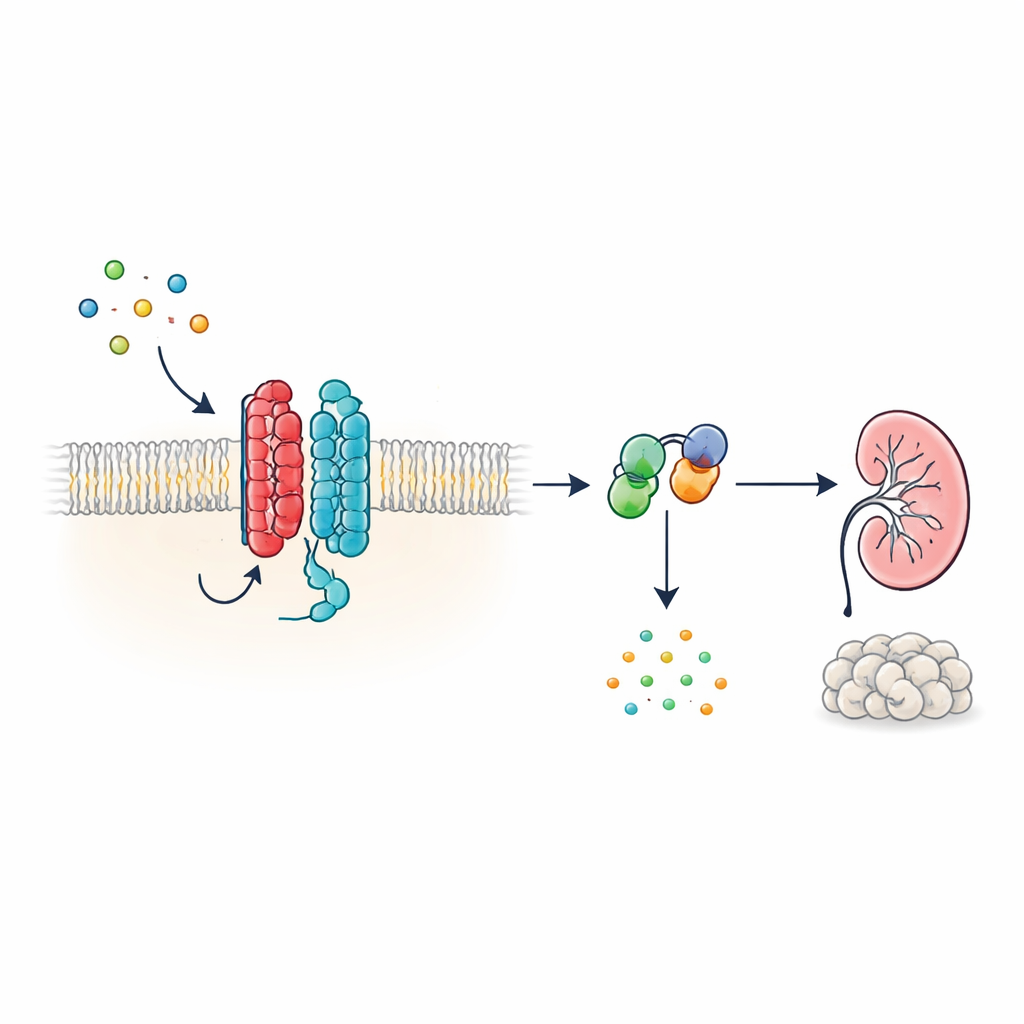
סוג חדש של אנטנה תאית
הורמונים ואותות רבים בגוף פועלים דרך קולטי חלבון‑G (GPCRs), משפחה עצומה של חלבוני ממברנה המכילים בדרך-כלל שבע מקטעים חוצי-ממברנה. PKD1, לעומת זאת, נראה שונה: יש לו 11 מקטעים והוא סווג יותר כערוץ יוני מאשר כקולטן קלאסי. הכותבים מראים שלמרות צורה בלתי שגרתית זו, PKD1 מתנהג כ‑GPCR אמיתי כאשר הוא בזוג עם PKD2. יחד, שני החלבונים יוצרים קומפלקס בממברנה שמגיב לחלבוני WNT—מולקולות סיגנל ורסטיליות המעורבות בהתפתחות ובתחזוקת הרקמות. כאשר WNT נקשרים ל‑PKD1, הקומפלקס מפעיל חלבונים‑G תוך-תאיים ומשנה רמות של cAMP, מולקולה קטנה המשפיעה רבות על גדילה תאית והפרשת נוזלים.
כיצד אותות WNT עוברים דרך קומפלקס PKD1–PKD2
כדי לעקוב אחר שרשרת הסיגנול הזו השתמשו החוקרים בחיישנים ביולוגיים מבוססי אור המדווחים על ארגון מחדש של חלבוני‑G בתאים חיים. בתאי מקור כלייתי אנושיים מהונדסים לבטא PKD1, מספר חברים במשפחת WNT גרמו לתת‑יחידות ספציפיות של חלבוני‑G להיפרד משותפיהן—מאפיין טיפוסי של הפעלה. התגובה הייתה חזקה ביותר לתת‑סוג חלבון‑G מעכב אחד, הידוע כ‑Gαi3, וכללה גם את Gαi1, Gαi2 ו‑Gαq הקשורים, אך לא את חלבוני‑G שלרוב מעלים cAMP. חיישנים נוספים הראו כי גירוי WNT לא רק קרב חלבוני‑G אל PKD1, אלא גם קידם את השלב הכימי בו GDP מוחלף ב‑GTP על תת‑היחידה Gα, ובכך השלים את מעגל ההפעלה. השפעות אלה הופיעו גם כאשר קולטני WNT ידועים אחרים הושתקו, מה שמעיד ש‑PKD1 עצמו, ולא קולטני WNT הקלאסיים, אחראי.
PKD2 כשותף חיוני ושומר השער
למרות ש‑PKD1 יכול לקשור גם ליגנדי WNT וגם חלבוני‑G, הוא אינו פועל לבדו. הצוות השתמש בעריכת גנום כדי להסיר את PKD2 מהתאים וגילה כי WNT כבר לא יכלה להניע הפעלת חלבון‑G דרך PKD1. החדרת PKD2 תקין שיחזרה את הסיגנל, אך וריאנטים של PKD2 הקשורים למחלה, שמפריעים להרכבת הקומפלקס או ליעדו לממברנה, לא שוחזרו. מיקרוסקופיה ומדידות העברת אנרגיה הראו כי PKD2 פועל כצ׳פרון, מלווה את PKD1 אל פני התא ומייצב אותו שם. חשוב לציין שמוטציות שמשנות את פעילות הערוץ היוני של PKD2 מבלי להשפיע על שותפותו עם PKD1 השאירו את סיגנל חלבוני‑G שלם, וחסימת זרימת יונים בעזרת מעכב כימי השפיעה מעט בלבד. זה מעיד כי, בהקשר זה, קומפלקס PKD1–PKD2 פועל בעיקר כקולטן ולא כערוץ יוני פעיל.
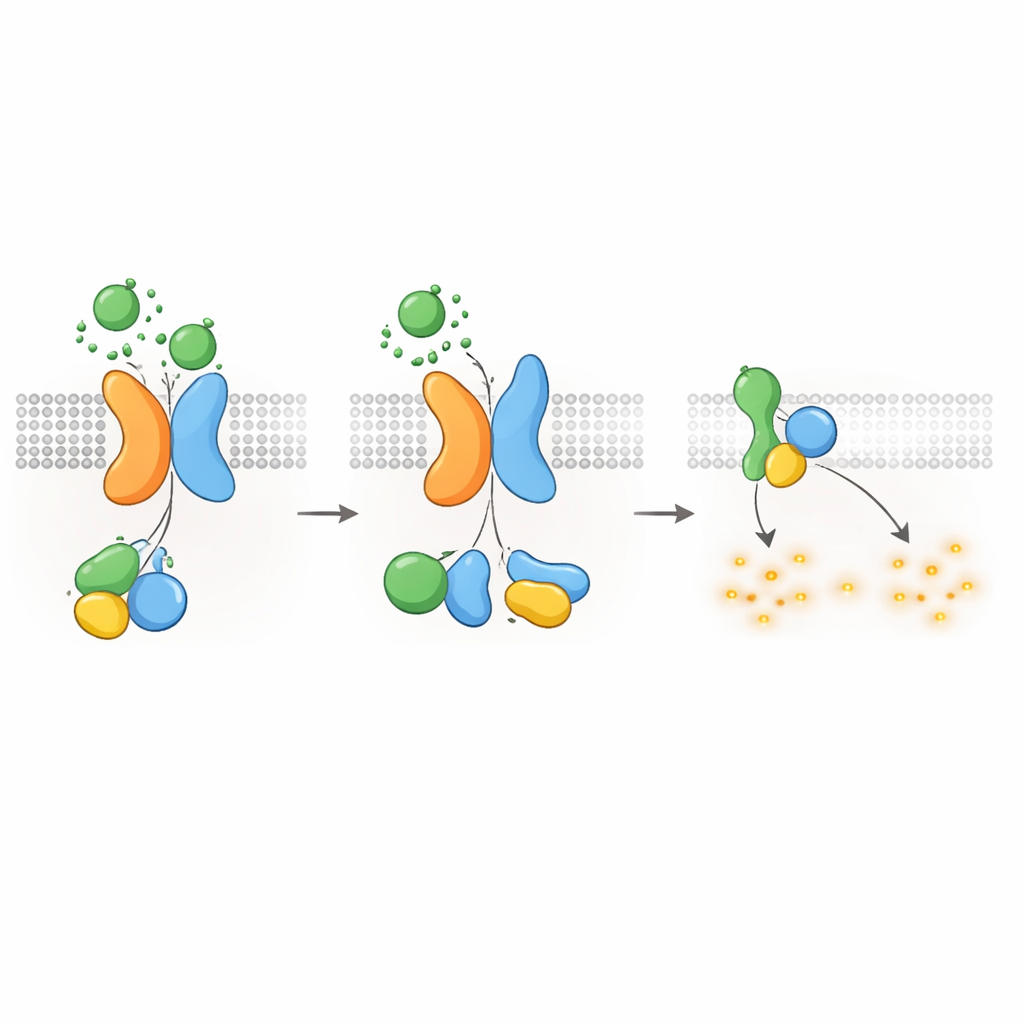
שמירה על cAMP—ויעיד ציסטות—תחת שליטה
מכיוון שחלבוני‑G מסוג Gαi ידועים בדיכוי יצירת cAMP, שאלו החוקרים האם קולט PKD1–PKD2 יכול להוריד ישירות רמות cAMP. באמצעות מדד זוהר שעוקב אחרי cAMP בזמן אמת הראו שהגירוי ב‑WNT בתאים מבטאים PKD1 האט את עליית ה‑cAMP שנגרמה על‑ידי פורסקולין, תרכובת שמגבירה יצירת cAMP. WNT גם הורידו רמות מנוחה של cAMP, והשניים נחסמו על‑ידי רעלן הפרטוסיס, שמנטרל ספציפית חלבוני Gαi. מוטציות ב‑PKD1 הגורמות מחלה—פוגעות בקישור ל‑WNT, בעיבוד אוטופרוטאוליטי או באינטראקציה עם חלבוני‑G—החלישו בחריפות את תגובת ההורדה של ה‑cAMP. תאים חסרי PKD2 גם לא הציגו דיכוי cAMP תלוי WNT, אלא אם הוחדר PKD2 תקין. ממצאים אלה מקשרים את הפגמים המולקולריים ב‑PKD1 או PKD2 ישירות לאובדן של הגבלת cAMP, מנגנון מרכזי בנהיגת גדילת הציסטות ב‑ADPKD.
כיבוי הסיגנל והעברתם של הקולטים פנימה
כמו ב‑GPCRs אחרים, קומפלקס PKD1–PKD2 חייב להיות מכובה ומחוזר לאחר ההפעלה. החוקרים גילו שמקינאז בשם GRK6, הנמצא בממברנת הפלסמה, מתקשר בחוזקה עם PKD1 כאשר PKD2 נוכח ועוזר בהסרת הקומפלקס מפני התא. חלבון נוסף, β‑arrestin 2, מגויס אז אל הקומפלקס המופעל ומלווה הן את PKD1 והן את PKD2 לאנדוזומים מוקדמים בתוך התא. חסימת פעילות GRK6 או פונקציית β‑arrestin הקטינה את הפניית הקומפלקס פנימה והשיבה חלקית את סיגנל חלבוני‑G. שלבי דה‑סנסיטיזציה אלה מרמזים שהקולטן PKD1–PKD2 מוסדר בקפידה, ונועדו למנוע דיכוי בלתי מבוקר של cAMP בתנאים רגילים.
מה משמעות הדבר למחלת כליה ומעבר לה
ביחד, התוצאות מקבעות את קומפלקס PKD1–PKD2 ככיתה חדשה של קולטן מצומד לחלבון‑G—קולטן לא שגרתי בעל 11 מקטעי טרנסממברנה התלוי בתת‑יחידה שותפה למיקום תקין על פני התא. בכליות בריאות, אותות WNT הפועלים דרך הקולטן הזה מפעילים חלבוני‑G מעכבים ושומרים על רמות cAMP בטווח בטוח, מסייעים לשמור על קוטר הצינורית הנורמלי ולמנוע היווצרות ציסטות. כאשר PKD1 או PKD2 מוטנטים, מסלול WNT–GPCR הזה נכשל, cAMP עולה והציסטות מתרחבות. על ידי הבהרת שלב ראשוני זה בתהליך המחלה, המחקר פותח דלתות לטיפולים שמטרתם לשקם או לחקות את הסיגנל של PKD1–PKD2, ובכך להציע חלופות מדויקות יותר לתרופות הנוכחיות שמורידות את ה‑cAMP באופן רחב יותר.
ציטוט: Hardy, E.P., Haider, A.N., Patel, M.M. et al. A heteromeric TRP channel that functions as a WNT-activated G protein-coupled receptor. Nat Commun 17, 3233 (2026). https://doi.org/10.1038/s41467-026-69932-w
מילות מפתח: מחלת כליות שחלתית פוליציסטית, PKD1 PKD2, קולטן מצומד לחלבון G, אותות WNT, cAMP