Clear Sky Science · nl
Een heteromere TRP-kanaal dat functioneert als een door WNT geactiveerde G-eiwitgekoppelde receptor
Waarom een nier-eiwit voor iedereen van belang is
Autosomal dominante polycysteuze nierziekte (ADPKD) is een van de meest voorkomende erfelijke nierziekten en een belangrijke oorzaak van nierfalen wereldwijd. Decennialang wisten wetenschappers welke genen de hoofdrol spelen—PKD1 en PKD2—but niet wat hun eiwitproducten precies in cellen doen. Dit artikel onthult dat deze eiwitten samen een ongebruikelijke receptor aan het celoppervlak vormen die rechtstreeks communiceert met klassieke schakelaars in de cel. Door uit te leggen hoe dit systeem normaal gesproken helpt een belangrijke boodschapperstof onder controle te houden, geeft het werk een duidelijker beeld van hoe cysten ontstaan bij ADPKD en wijst het op nieuwe behandelrichtingen.
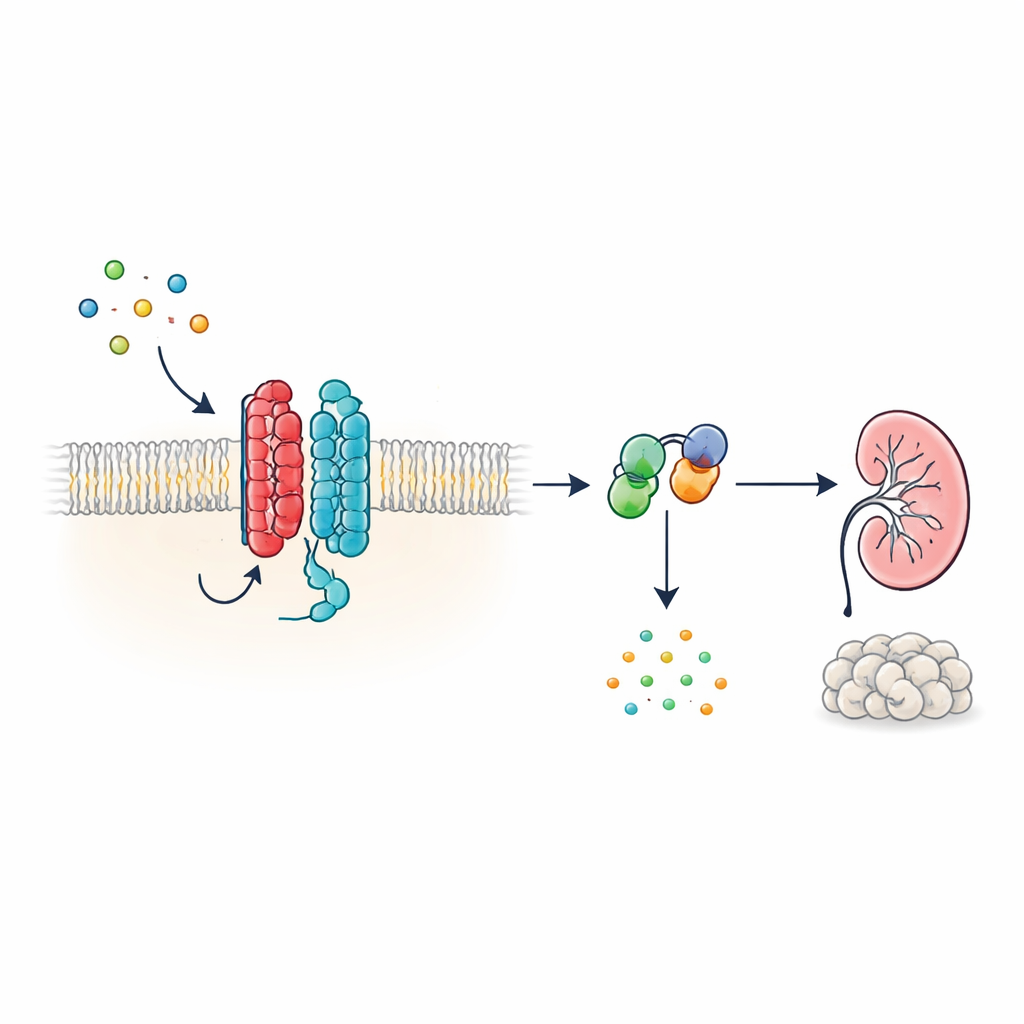
Een nieuw type cellulair antenne
Veel hormonen en signalen in het lichaam werken via G-eiwitgekoppelde receptoren (GPCRs), een enorme familie van membraan-eiwitten met een kenmerkende zevensegmentstructuur die door het celmembraan loopt. PKD1 ziet er echter anders uit: het heeft 11 segmenten en is eerder geclassificeerd bij ionkanalen dan bij klassieke receptoren. De auteurs tonen aan dat PKD1, ondanks deze onconventionele vorm, zich als een volwaardige GPCR gedraagt wanneer het gepaard gaat met PKD2. Samen vormen de twee eiwitten een complex in het membraan dat reageert op WNT-eiwitten—veelzijdige signaalmoleculen betrokken bij ontwikkeling en weefselonderhoud. Wanneer WNTs aan PKD1 binden, activeert het complex intracellulaire G-eiwitten en verandert het de niveaus van cyclisch AMP (cAMP), een klein molecuul dat sterk van invloed is op celgroei en vloeistofsecretie.
Hoe WNT-signalen door het PKD1–PKD2-complex reizen
Om deze signaalketen te volgen gebruikten de onderzoekers lichtgebaseerde biosensoren die rapporteren hoe G-eiwitten zich herschikken in levende cellen. In menselijk nier-afgeleide cellen die waren aangepast om PKD1 tot expressie te brengen veroorzaakten meerdere verschillende WNT-leden dat specifieke G-eiwitsubunits zich van hun partners afscheidden—een kenmerk van activatie. De respons was het sterkst voor een remmend G-eiwitsubtype, bekend als Gαi3, en betrof ook verwante Gαi1-, Gαi2- en Gαq-subunits, maar niet de G-eiwitten die doorgaans cAMP verhogen. Aanvullende biosensoren toonden aan dat WNT-stimulatie niet alleen G-eiwitten in de nabijheid van PKD1 bracht, maar ook de chemische stap aandreef waarbij GDP voor GTP op de Gα-subunit wordt verwisseld, waarmee de activatiecyclus wordt voltooid. Deze effecten traden op zelfs wanneer andere bekende WNT-receptoren werden onderdrukt, wat aangeeft dat PKD1 zelf, in plaats van klassieke WNT-receptoren, verantwoordelijk was.
PKD2 als essentiële partner en poortwachter
Hoewel PKD1 zowel WNT-liganden als G-eiwitten kan binden, staat het niet alleen. Het team gebruikte genoombewerking om PKD2 uit cellen te verwijderen en ontdekte dat WNT niet langer G-eiwitactivatie via PKD1 kon aandrijven. Het opnieuw inbrengen van normaal PKD2 herstelde de signaaltransductie, maar ziekte-geassocieerde PKD2-varianten die de complexassemblage of gerichte membraanlocatie verstoren deden dat niet. Microscopia en energieoverdrachtsmetingen toonden aan dat PKD2 fungeert als chaperonne, PKD1 naar het celoppervlak begeleidt en daar stabiliseert. Belangrijk is dat mutaties die de ionkanaalactiviteit van PKD2 veranderen zonder de samenwerking met PKD1 aan te tasten, de G-eiwitsignalering ongemoeid lieten, en het blokkeren van ionstroom met een chemische remmer weinig effect had. Dit duidt erop dat het PKD1–PKD2-complex in deze context hoofdzakelijk functioneert als receptor in plaats van als actief ionkanaal.
cAMP—en cystengroei—onder controle houden
Aangezien Gαi-type G-eiwitten bekendstaan om het dempen van cAMP-productie, vroegen de auteurs zich vervolgens af of de PKD1–PKD2-receptor cAMP-niveaus direct naar beneden kon brengen. Met een luminescente reporter die cAMP in realtime volgt, toonden zij aan dat WNT-stimulatie in PKD1-expressiecellen de stijging van cAMP, veroorzaakt door forskolin (een stof die cAMP-productie verhoogt), vertraagde. WNTs verlaagden ook de rustniveaus van cAMP, en beide effecten werden geblokkeerd door pertussis-toxine, dat specifiek Gαi-eiwitten uitschakelt. Ziekte-veroorzakende PKD1-mutaties—die WNT-binding, autoproteolytische verwerking of G-eiwitinteractie aantasten—verzwakten deze cAMP-verlagende respons sterk. Cellen zonder PKD2 toonden eveneens geen WNT-afhankelijke cAMP-suppressie, tenzij normaal PKD2 werd heringevoerd. Deze bevindingen koppelen de moleculaire defecten in PKD1 of PKD2 rechtstreeks aan verlies van cAMP-remming, een centrale drijfkracht van cystengroei bij ADPKD.
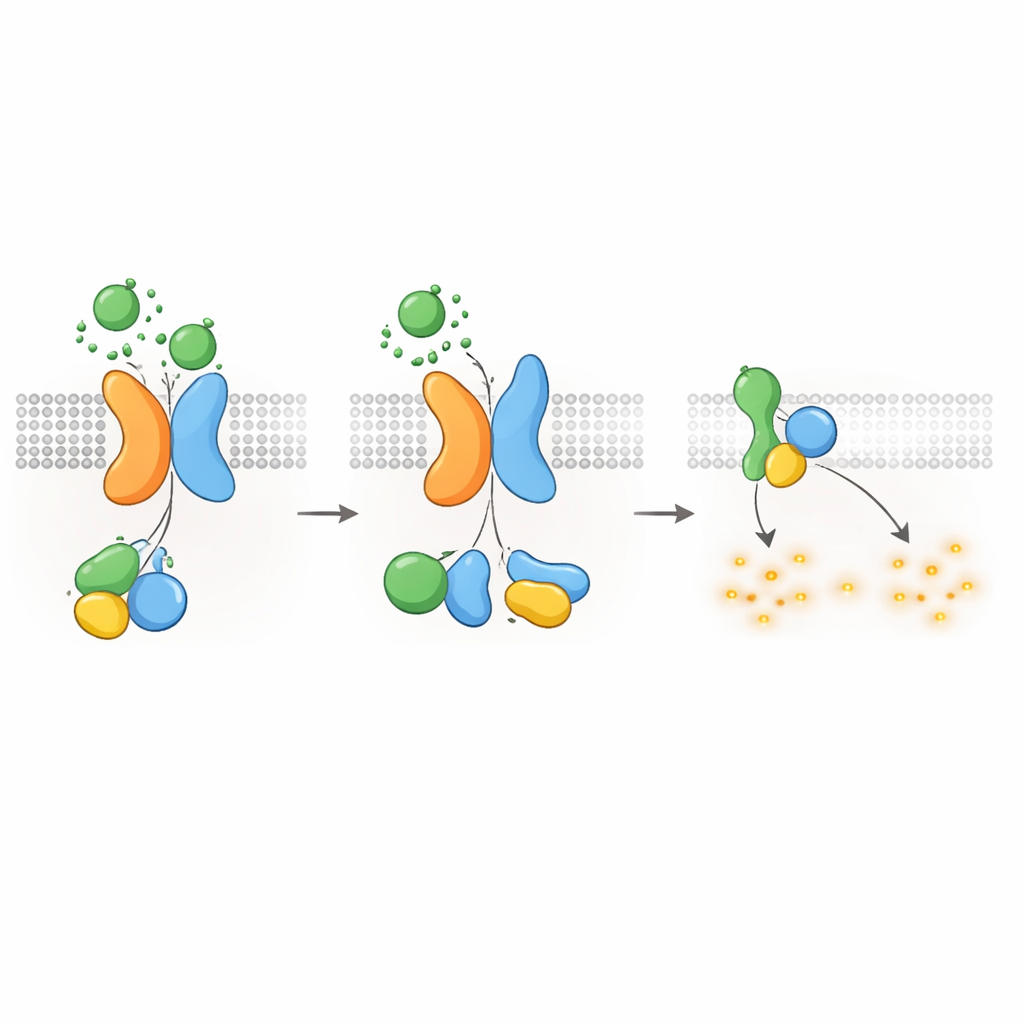
Het signaal uitschakelen en receptoren naar binnen verplaatsen
Net als andere GPCRs moet het PKD1–PKD2-complex na activatie worden uitgeschakeld en gerecycleerd. De auteurs vonden dat een kinase genaamd GRK6, die zich aan het plasmamembraan bevindt, sterk met PKD1 geassocieerd is wanneer PKD2 aanwezig is en helpt het complex van het celoppervlak te verwijderen. Een ander eiwit, β-arrestine 2, wordt vervolgens naar het geactiveerde complex gerekruteerd en begeleidt zowel PKD1 als PKD2 naar vroege endosomen in de cel. Het blokkeren van GRK6-activiteit of β-arrestinefunctie verminderde deze internalisatie en herstelde gedeeltelijk de G-eiwitsignalering. Deze desensitisatiestappen suggereren dat de PKD1–PKD2-receptor strak gereguleerd is, waardoor een doorgeschoten onderdrukking van cAMP onder normale omstandigheden wordt voorkomen.
Wat dit betekent voor nierziekten en verder
Samen laten de resultaten zien dat het PKD1–PKD2-complex een nieuwe klasse van G-eiwitgekoppelde receptor vormt—een onconventionele 11-transmembraanreceptor die afhankelijk is van een partner-subunit voor correcte plaatsing aan het celoppervlak. In gezonde nieren activeren WNT-signalen via deze receptor remmende G-eiwitten en houden ze cAMP-niveaus binnen veilige grenzen, wat helpt om normale buisdiameters te behouden en cystevorming te voorkomen. Wanneer PKD1 of PKD2 gemuteerd raakt, hapert deze WNT–GPCR-route, stijgt cAMP en breiden cysten zich uit. Door deze vroege stap in het ziekteproces te verduidelijken, opent de studie de deur naar therapieën gericht op het herstellen of nabootsen van PKD1–PKD2-signaaltransductie, wat mogelijk preciezere alternatieven biedt voor huidige geneesmiddelen die cAMP meer algemeen dempen.
Bronvermelding: Hardy, E.P., Haider, A.N., Patel, M.M. et al. A heteromeric TRP channel that functions as a WNT-activated G protein-coupled receptor. Nat Commun 17, 3233 (2026). https://doi.org/10.1038/s41467-026-69932-w
Trefwoorden: polycysteuze nierziekte, PKD1 PKD2, G-eiwitgekoppelde receptor, WNT-signaaltransductie, cAMP