Clear Sky Science · es
Un canal TRP heteromérico que funciona como un receptor acoplado a proteína G activado por WNT
Por qué una proteína renal importa a todo el mundo
La enfermedad renal poliquística autosómica dominante (ERPAD) es uno de los trastornos renales hereditarios más comunes y una causa principal de insuficiencia renal en todo el mundo. Durante décadas, los científicos han identificado los genes principales implicados—PKD1 y PKD2—pero no sabían qué hacen exactamente sus productos proteicos dentro de las células. Este artículo revela que estas proteínas forman un receptor inusual en la superficie celular que comunica directamente con los interruptores clásicos de señalización intracelular. Al explicar cómo este sistema normalmente ayuda a mantener controlada una molécula mensajera clave, el trabajo ofrece una imagen más nítida de cómo se originan los quistes en la ERPAD y apunta a nuevas vías para tratar la enfermedad.
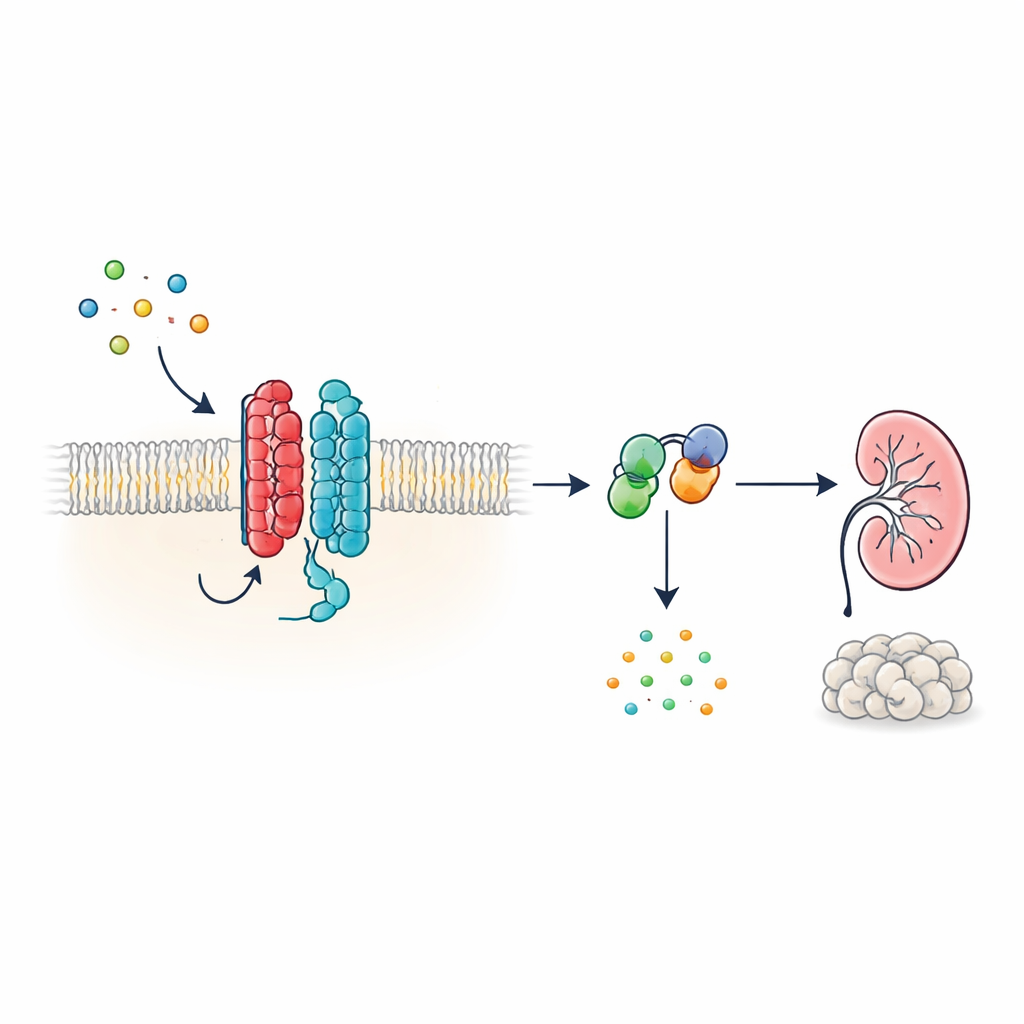
Un nuevo tipo de antena celular
Muchas hormonas y señales en el organismo actúan a través de receptores acoplados a proteína G (GPCR), una gran familia de proteínas de superficie celular con una estructura característica de siete segmentos que atraviesa la membrana. PKD1, sin embargo, tiene un aspecto diferente: presenta 11 segmentos y se ha clasificado más con canales iónicos que con receptores clásicos. Los autores muestran que, pese a esta forma poco convencional, PKD1 se comporta como un verdadero GPCR cuando se asocia con PKD2. Juntas, las dos proteínas forman un complejo en la membrana que responde a las proteínas WNT—moléculas de señalización versátiles implicadas en el desarrollo y el mantenimiento tisular. Cuando las WNT se unen a PKD1, el complejo activa proteínas G intracelulares y modifica los niveles de AMP cíclico (cAMP), una pequeña molécula que influye de forma potente en el crecimiento celular y la secreción de líquido.
Cómo viajan las señales WNT a través del complejo PKD1–PKD2
Para trazar esta cadena de señalización, los investigadores utilizaron biosensores basados en luz que informan cómo se reorganizan las proteínas G en células vivas. En células humanas derivadas del riñón diseñadas para expresar PKD1, varios miembros de la familia WNT provocaron que distintos subunidades de proteínas G se separaran de sus parejas—una señal distintiva de activación. La respuesta fue más intensa para una subfamilia inhibitoria de proteína G, conocida como Gαi3, e implicó también a las subunidades relacionadas Gαi1, Gαi2 y Gαq, pero no a las proteínas G que típicamente aumentan el cAMP. Biosensores adicionales mostraron que la estimulación por WNT no solo acercó las proteínas G a PKD1, sino que también impulsó el paso químico en el que el GDP se intercambia por GTP en la subunidad Gα, completando el ciclo de activación. Estos efectos ocurrieron incluso cuando se suprimieron otros receptores WNT conocidos, lo que indica que fue el propio PKD1, más que los receptores WNT clásicos, el responsable.
PKD2 como socio esencial y guardián
Aunque PKD1 puede unirse tanto a ligandos WNT como a proteínas G, no actúa en solitario. El equipo utilizó edición genómica para eliminar PKD2 de las células y descubrió que WNT ya no podía inducir la activación de proteínas G a través de PKD1. La reintroducción de PKD2 normal restauró la señalización, pero variantes de PKD2 asociadas a la enfermedad que interfieren en el ensamblaje del complejo o en la localización en membrana no lo hicieron. La microscopía y las mediciones de transferencia de energía mostraron que PKD2 actúa como un chaperón, escoltando a PKD1 hasta la superficie celular y estabilizándolo allí. De forma importante, mutaciones que alteran la actividad de canal iónico de PKD2 sin afectar su asociación con PKD1 dejaron intacta la señalización por proteínas G, y bloquear el flujo iónico con un inhibidor químico tuvo poco efecto. Esto indica que, en este contexto, el complejo PKD1–PKD2 funciona principalmente como un receptor y no tanto como un canal iónico operativo.
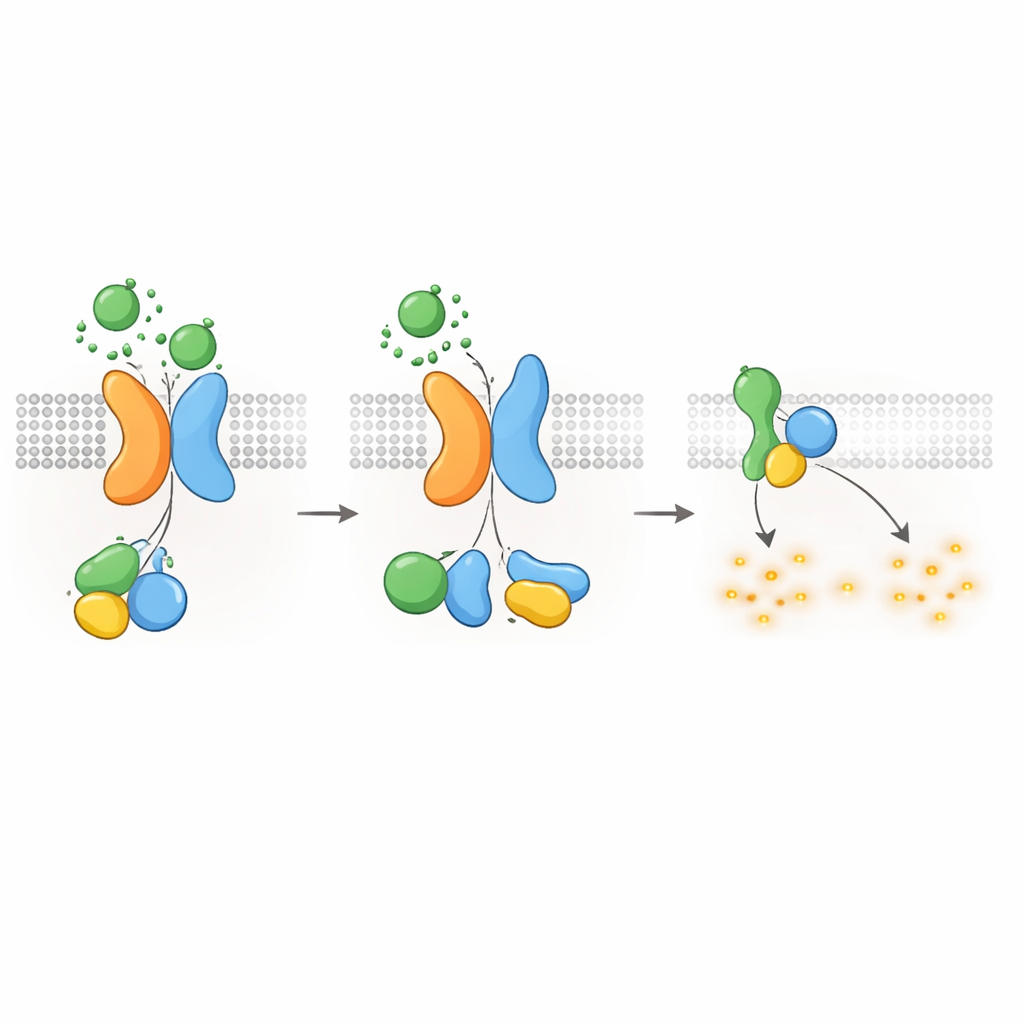
Mantener el cAMP—y el crecimiento de quistes—bajo control
Dado que se sabe que las proteínas G del tipo Gαi reducen la producción de cAMP, los autores preguntaron si el receptor PKD1–PKD2 podía disminuir directamente los niveles de cAMP. Usando un reportero luminiscente que sigue el cAMP en tiempo real, mostraron que la estimulación por WNT en células que expresan PKD1 ralentizaba el aumento de cAMP provocado por la forskolina, un compuesto que potencia la producción de cAMP. Las WNT también redujeron los niveles basales de cAMP, y ambos efectos fueron bloqueados por la toxina pertussis, que inactiva específicamente las proteínas Gαi. Mutaciones patogénicas en PKD1—que afectan la unión a WNT, el procesamiento autoproteolítico o la interacción con proteínas G—debilitaron de forma marcada esta respuesta reductora de cAMP. Las células carentes de PKD2 tampoco mostraron supresión dependiente de WNT del cAMP, a menos que se reintrodujera PKD2 normal. Estos hallazgos conectan los defectos moleculares en PKD1 o PKD2 directamente con la pérdida del control del cAMP, un impulsor central del crecimiento quístico en la ERPAD.
Apagar la señal y mover los receptores al interior
Como otros GPCR, el complejo PKD1–PKD2 debe apagarse y reciclarse tras la activación. Los autores hallaron que una quinasa llamada GRK6, que reside en la membrana plasmática, se asocia fuertemente con PKD1 cuando PKD2 está presente y ayuda a retirar el complejo de la superficie celular. Otra proteína, la β-arrestina 2, es entonces reclutada al complejo activado y escolta tanto a PKD1 como a PKD2 hacia endosomas tempranos dentro de la célula. Bloquear la actividad de GRK6 o la función de la β-arrestina redujo esta internalización y restauró parcialmente la señalización por proteínas G. Estos pasos de desensibilización sugieren que el receptor PKD1–PKD2 está fuertemente regulado, evitando una supresión descontrolada del cAMP en condiciones normales.
Qué significa esto para la enfermedad renal y más allá
En conjunto, los resultados establecen al complejo PKD1–PKD2 como una nueva clase de receptor acoplado a proteína G—un receptor no convencional de 11 transmembranas que depende de una subunidad asociada para su correcta localización en la superficie celular. En riñones sanos, las señales WNT que actúan a través de este receptor activan proteínas G inhibitorias y mantienen los niveles de cAMP dentro de un rango seguro, ayudando a conservar el diámetro tubular normal y a prevenir la formación de quistes. Cuando PKD1 o PKD2 están mutados, esta vía WNT–GPCR falla, el cAMP aumenta y los quistes se expanden. Al aclarar este paso temprano en el proceso de la enfermedad, el estudio abre la puerta a terapias dirigidas a restaurar o imitar la señalización PKD1–PKD2, ofreciendo potencialmente alternativas más precisas a los fármacos actuales que reducen el cAMP de forma más amplia.
Cita: Hardy, E.P., Haider, A.N., Patel, M.M. et al. A heteromeric TRP channel that functions as a WNT-activated G protein-coupled receptor. Nat Commun 17, 3233 (2026). https://doi.org/10.1038/s41467-026-69932-w
Palabras clave: enfermedad renal poliquística, PKD1 PKD2, receptor acoplado a proteína G, señalización WNT, cAMP