Clear Sky Science · ru
Гетеромерный канал TRP, функционирующий как активируемый WNT G-белок-связывающий рецептор
Почему белок почки важен для всех
Аутосомно-доминантный поликистоз почек (ADPKD) — одно из самых распространённых наследственных заболеваний почек и одна из ведущих причин почечной недостаточности в мире. Долгое время учёным были известны ключевые гены — PKD1 и PKD2 — но оставалось неясным, что именно делают их белки в клетке. В этой статье показано, что эти белки формируют необычный рецептор на поверхности клетки, который напрямую взаимодействует с классическими сигнальными переключателями внутри клетки. Объясняя, как эта система обычно сдерживает ключевой посредник сигнала, работа даёт более ясное представление о том, как образуются кисты при ADPKD, и указывает на новые подходы к лечению болезни.
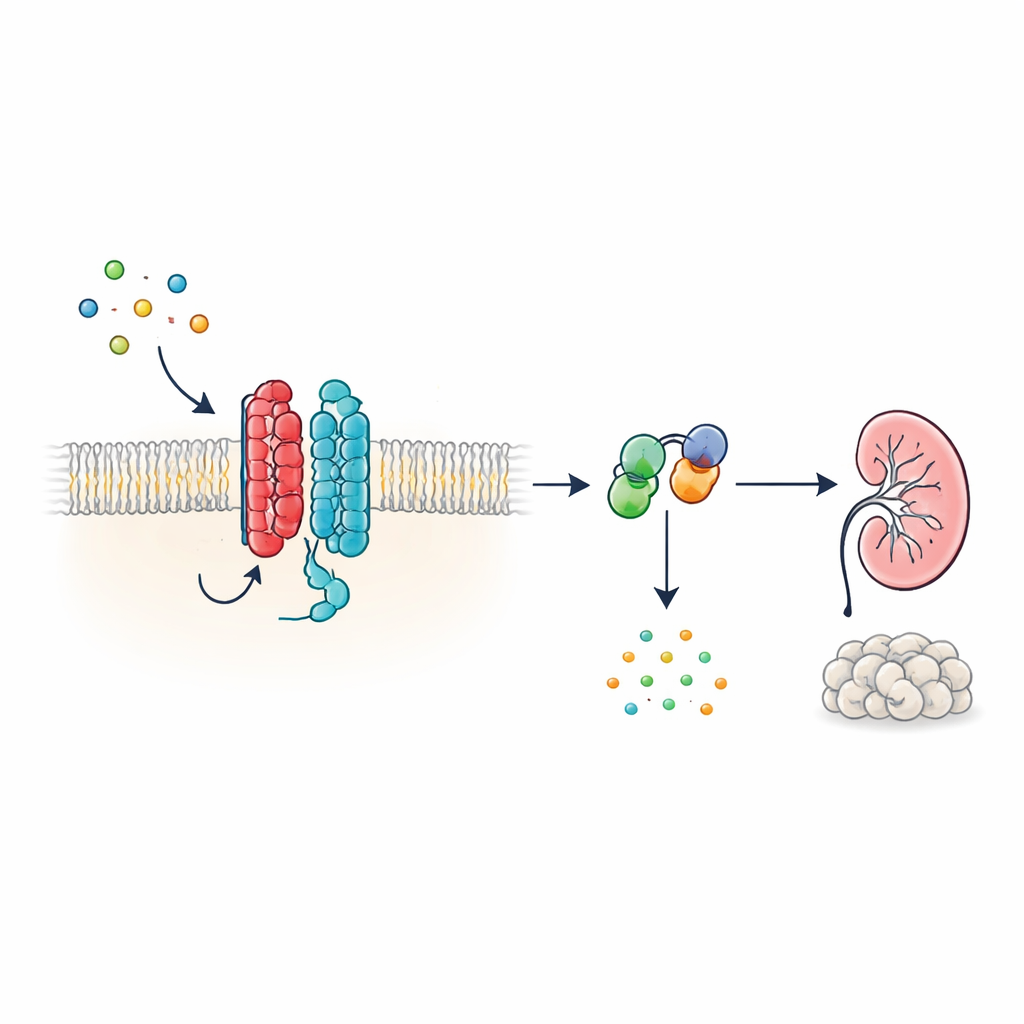
Новый тип клеточной антенны
Многие гормоны и сигналы в организме действуют через G-белок-связывающие рецепторы (GPCR), большую семью белков на поверхности клетки с характерной семисегментной структурой, проходящей через мембрану. PKD1 выглядит иначе: у него 11 трансмембранных сегментов, и его раньше относили к ионным каналам, а не к классическим рецепторам. Авторы показывают, что несмотря на такую необычную форму, PKD1 ведёт себя как настоящий GPCR в сочетании с PKD2. Вместе эти два белка формируют комплекс в мембране, который отвечает на белки WNT — универсальные сигнальные молекулы, участвующие в развитии и поддержании тканей. При связывании WNT с PKD1 комплекс активирует внутриклеточные G‑белки и изменяет уровни циклического АМФ (цАМФ), небольшого молекулы, сильно влияющей на рост клеток и секрецию жидкости.
Как WNT-сигнал проходит через комплекс PKD1–PKD2
Чтобы проследить эту сигнальную цепочку, исследователи использовали оптические биосенсоры, которые показывают, как G‑белки перестраиваются в живых клетках. В клетках, происходящих из человеческой почки и генетически запрограммированных на экспрессию PKD1, несколько членов семейства WNT вызывали расщепление отдельных субъединиц G‑белков от их партнёров — признак активации. Наиболее сильный ответ отмечался для одного ингибирующего подкласса G‑белков, известного как Gαi3, а также вовлекались родственные субъединицы Gαi1, Gαi2 и Gαq, но не те G‑белки, которые обычно повышают цАМФ. Дополнительные биосенсоры показали, что стимуляция WNT не только сближает G‑белки с PKD1, но и вызывает химический шаг замены GDP на GTP на субъединице Gα, завершая цикл активации. Эти эффекты наблюдались даже при подавлении других известных WNT‑рецепторов, что указывает на то, что именно PKD1, а не классические WNT‑рецепторы, отвечает за сигнализацию.
PKD2 как необходимый партнёр и страж
Хотя PKD1 способен связывать как лиганды WNT, так и G‑белки, он не действует в одиночку. Команда использовала редактирование генома для удаления PKD2 из клеток и обнаружила, что WNT больше не может вызывать активацию G‑белков через PKD1. Повторное введение нормального PKD2 восстановило сигнализацию, тогда как варианты PKD2, связанные с болезнью и нарушающие сборку комплекса или направленность в мембрану, этого не сделали. Микроскопия и измерения переноса энергии показали, что PKD2 действует как шаперон, сопровождая PKD1 на поверхность клетки и стабилизируя его там. Важно, что мутации, изменяющие ионную активность PKD2, но не влияющие на его партнёрство с PKD1, не нарушали G‑белковую сигнализацию, а блокирование потока ионов химическим ингибитором имело незначительный эффект. Это указывает на то, что в данном контексте комплекс PKD1–PKD2 в первую очередь функционирует как рецептор, а не как работающий ионный канал.
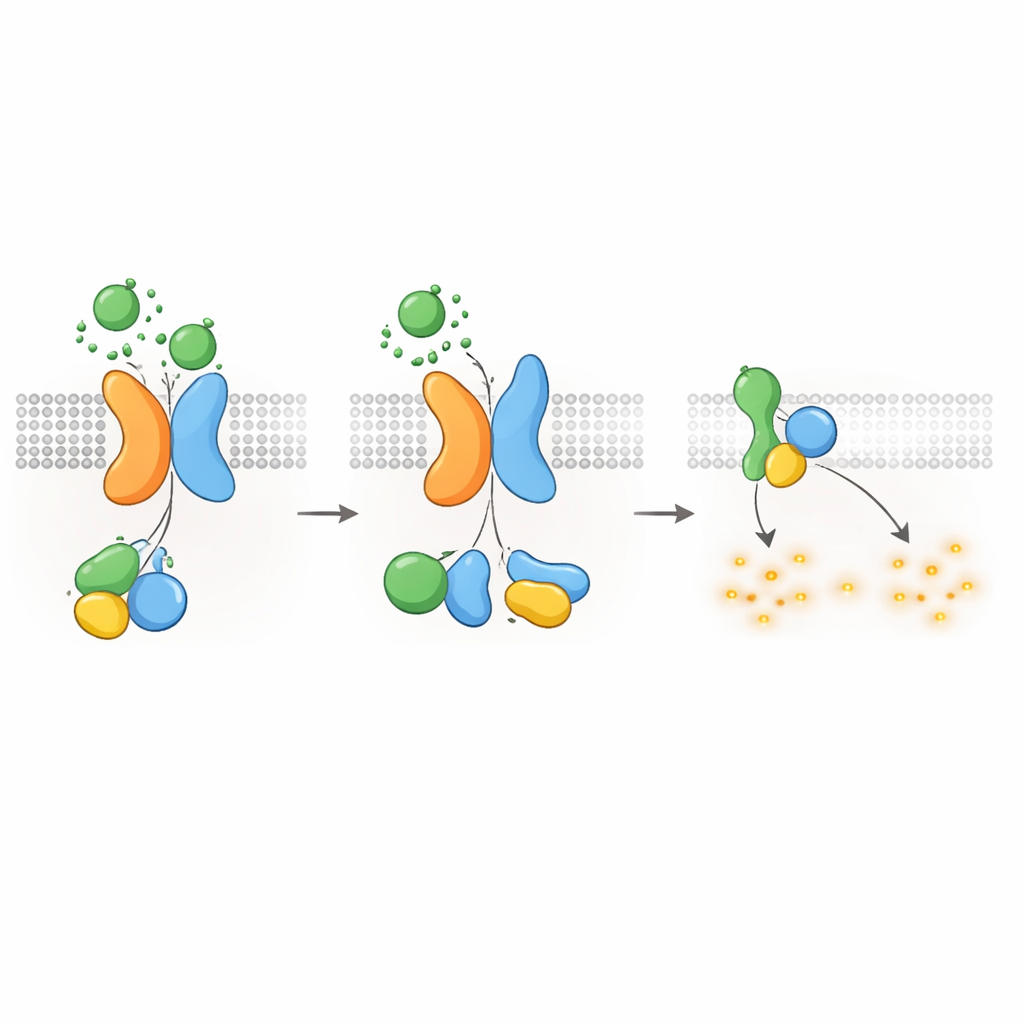
Сдерживание цАМФ — и рост кист
Поскольку известно, что G‑белки типа Gαi подавляют продукцию цАМФ, авторы проверили, может ли рецептор PKD1–PKD2 непосредственно понижать уровни цАМФ. Используя люминесцентный репортёр для отслеживания цАМФ в реальном времени, они показали, что стимуляция WNT в клетках, экспрессирующих PKD1, замедляла повышение цАМФ, вызванное форсколином, соединением, усиливающим продукцию цАМФ. WNT также снижали уровень базального цАМФ, и оба эффекта блокировались токсоидом, который специфически отключает белки Gαi. Болезнетворные мутации PKD1 — затрагивающие связывание WNT, автопротеолитическую обработку или взаимодействие с G‑белками — существенно ослабляли этот ответ на понижение цАМФ. Клетки без PKD2 также не проявляли подавления цАМФ при стимуляции WNT, если только в них заново не ввести нормальный PKD2. Эти результаты связывают молекулярные дефекты в PKD1 или PKD2 напрямую с потерей контроля над цАМФ, что является ключевым фактором роста кист при ADPKD.
Выключение сигнала и перемещение рецепторов внутрь клетки
Как и у других GPCR, комплекс PKD1–PKD2 должен выключаться и перерабатываться после активации. Авторы обнаружили, что киназа GRK6, локализующаяся в плазматической мембране, прочно ассоциируется с PKD1 в присутствии PKD2 и способствует удалению комплекса с поверхности клетки. Затем к активированному комплексу привлекается белок β‑аррестин 2, который сопровождает и PKD1, и PKD2 в ранние эндосомы внутри клетки. Блокирование активности GRK6 или функции β‑аррестина уменьшало эту интернализацию и частично восстанавливало G‑белковую сигнализацию. Эти шаги десенситизации указывают на то, что рецептор PKD1–PKD2 строго регулируется, предотвращая чрезмерное подавление цАМФ в нормальных условиях.
Что это значит для заболеваний почек и не только
В целом результаты устанавливают комплекс PKD1–PKD2 как новый класс G‑белок‑связывающего рецептора — необычный 11‑трансмембранный рецептор, зависящий от партнёрной субъединицы для правильного расположения на поверхности клетки. В здоровых почках WNT‑сигналы, действующие через этот рецептор, активируют ингибирующие G‑белки и поддерживают уровни цАМФ в безопасном диапазоне, помогая сохранять нормальный диаметр канальцев и предотвращать образование кист. Когда PKD1 или PKD2 мутируют, этот путь WNT–GPCR даёт сбой, цАМФ повышается, и кисты растут. Проясняя этот ранний этап процесса болезни, исследование открывает путь к терапиям, направленным на восстановление или имитацию сигнализации PKD1–PKD2, что потенциально может предложить более точные альтернативы текущим препаратам, которые в более широком виде снижают цАМФ.
Цитирование: Hardy, E.P., Haider, A.N., Patel, M.M. et al. A heteromeric TRP channel that functions as a WNT-activated G protein-coupled receptor. Nat Commun 17, 3233 (2026). https://doi.org/10.1038/s41467-026-69932-w
Ключевые слова: поликистоз почек, PKD1 PKD2, G-белок-связывающий рецептор, WNT-сигнализация, цАМФ