Clear Sky Science · it
Un canale TRP eteromerico che funziona come un recettore accoppiato a proteine G attivato da WNT
Perché una proteina renale conta per tutti
La malattia renale policistica autosomica dominante (ADPKD) è uno dei disordini renali ereditari più comuni ed è una causa principale di insufficienza renale in tutto il mondo. Per decenni gli scienziati hanno conosciuto i geni principali coinvolti—PKD1 e PKD2—ma non cosa facessero effettivamente i loro prodotti proteici all’interno delle cellule. Questo lavoro rivela che queste proteine formano un recettore insolito sulla superficie cellulare che comunica direttamente con i classici interruttori di segnalazione intracellulari. Spiegando come questo sistema aiuta normalmente a mantenere sotto controllo un messaggero chiave, lo studio offre un quadro più chiaro di come si formino le cisti nell’ADPKD e indica nuove possibili strade terapeutiche.
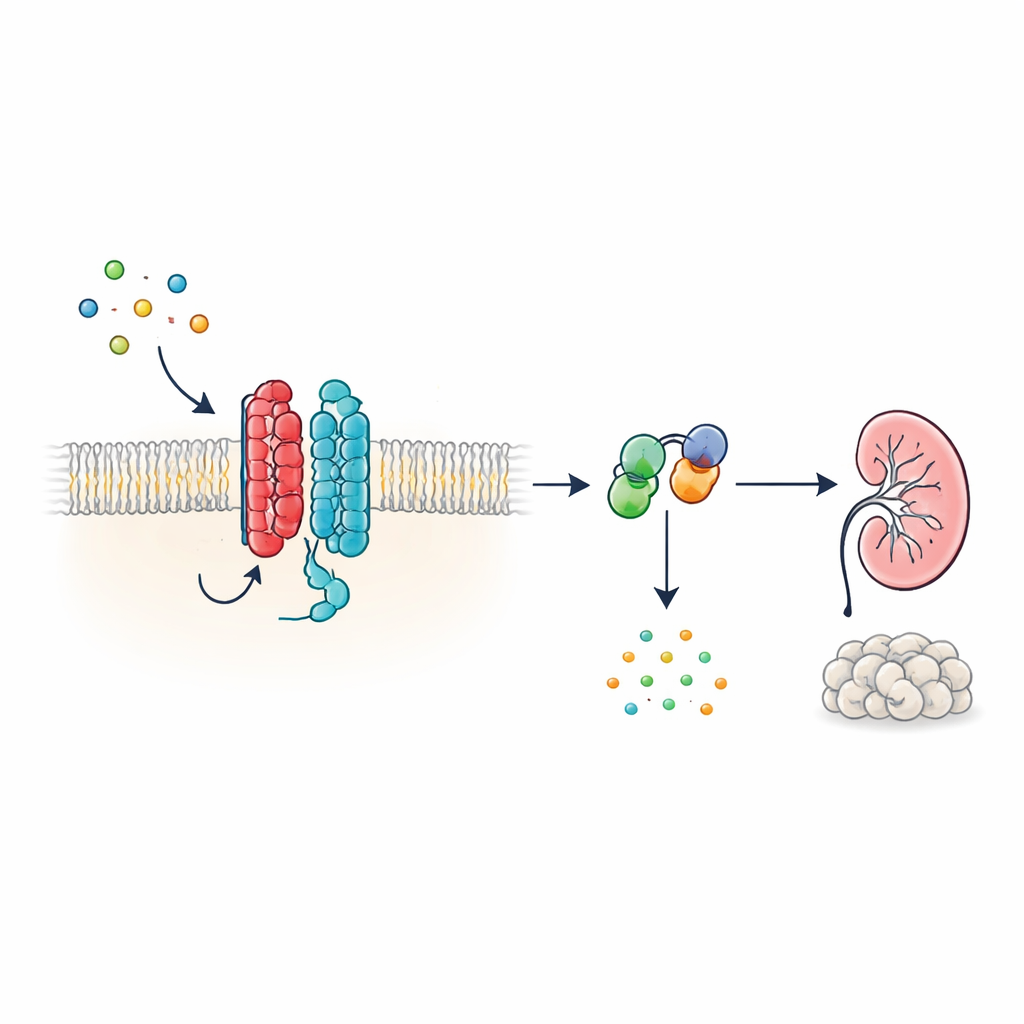
Un nuovo tipo di antenna cellulare
Molti ormoni e segnali nell’organismo agiscono tramite recettori accoppiati a proteine G (GPCR), una grande famiglia di proteine di superficie con una caratteristica struttura a sette eliche che attraversano la membrana cellulare. PKD1, tuttavia, appare diversa: possiede 11 eliche transmembrana ed è stata classificata più tra i canali ionici che tra i recettori classici. Gli autori mostrano che, nonostante questa forma non convenzionale, PKD1 si comporta come un vero GPCR quando è associata a PKD2. Insieme, le due proteine formano un complesso nella membrana che risponde alle proteine WNT—molecole di segnalazione versatili coinvolte nello sviluppo e nel mantenimento dei tessuti. Quando le WNT si legano a PKD1, il complesso attiva le proteine G intracellulari e modifica i livelli di cAMP (AMP ciclico), una piccola molecola che influenza fortemente la crescita cellulare e la secrezione di fluidi.
Come i segnali WNT viaggiano attraverso il complesso PKD1–PKD2
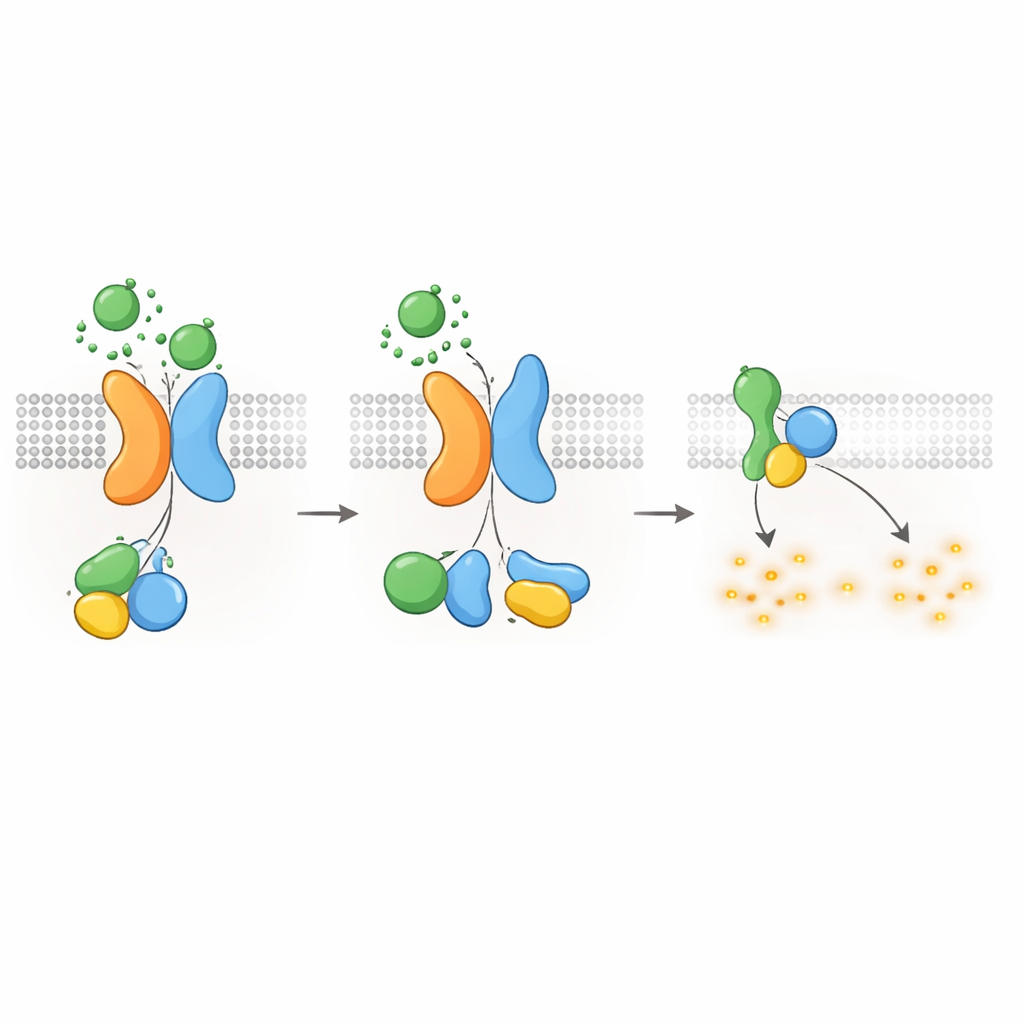
Per tracciare questa catena di segnalazione, i ricercatori hanno utilizzato biosensori ottici che riportano come le proteine G si riorganizzano nelle cellule viventi. In cellule umane derivate dal rene, ingegnerizzate per esprimere PKD1, diversi membri della famiglia WNT hanno fatto sì che specifici sottounità delle proteine G si separassero dai loro partner—un segno caratteristico di attivazione. La risposta è stata più forte per un sottotipo inibitorio di proteina G, noto come Gαi3, e ha coinvolto anche Gαi1, Gαi2 e sottounità Gαq correlate, ma non le proteine G che tipicamente aumentano il cAMP. Biosensori aggiuntivi hanno mostrato che la stimolazione WNT non solo avvicinava le proteine G a PKD1, ma promuoveva anche il passaggio chimico in cui il GDP viene scambiato con il GTP sulla subunità Gα, completando il ciclo di attivazione. Questi effetti si sono verificati anche quando altri recettori WNT noti erano soppressi, indicando che è PKD1 stesso, piuttosto che i recettori WNT classici, il responsabile.
PKD2 come partner essenziale e guardiano
Nonostante PKD1 possa legare sia i ligandi WNT sia le proteine G, non agisce da sola. Il team ha usato l’editing genomico per rimuovere PKD2 dalle cellule e ha constatato che WNT non riusciva più a promuovere l’attivazione delle proteine G tramite PKD1. Reintrodurre PKD2 normale ha restaurato la segnalazione, ma varianti patologiche di PKD2 che interrompono l’assemblaggio del complesso o il targeting alla membrana non l’hanno fatto. Microscopia e misure di trasferimento di energia hanno mostrato che PKD2 agisce come una chaperona, escortando PKD1 alla superficie cellulare e stabilizzandolo lì. È importante notare che mutazioni che alterano l’attività di canale ionico di PKD2 senza compromettere la sua associazione con PKD1 hanno lasciato intatta la segnalazione delle proteine G, e bloccare il flusso ionico con un inibitore chimico ha avuto scarso effetto. Ciò indica che, in questo contesto, il complesso PKD1–PKD2 funziona principalmente come recettore piuttosto che come un canale ionico operativo.
Mantenere sotto controllo il cAMP—e la crescita delle cisti
Poiché le proteine G di tipo Gαi sono note per attenuare la produzione di cAMP, gli autori si sono chiesti se il recettore PKD1–PKD2 potesse abbassare direttamente i livelli di cAMP. Usando un reporter luminescente che monitora il cAMP in tempo reale, hanno mostrato che la stimolazione WNT in cellule che esprimevano PKD1 rallentava l’aumento di cAMP innescato dalla forskolina, un composto che incrementa la produzione di cAMP. Le WNT abbassavano anche i livelli basali di cAMP, e entrambi gli effetti venivano bloccati dalla tossina pertussica, che disabilita specificamente le proteine Gαi. Mutazioni patogene in PKD1—che compromettono il legame con WNT, il processamento autoproteolitico o l’interazione con le proteine G—indebolivano drasticamente questa risposta di riduzione del cAMP. Anche le cellule prive di PKD2 non mostravano la soppressione del cAMP dipendente da WNT, a meno che non venisse reintrodotta PKD2 normale. Questi risultati collegano i difetti molecolari in PKD1 o PKD2 direttamente alla perdita del controllo del cAMP, un motore centrale della crescita cistica nell’ADPKD.
Spegnere il segnale e spostare i recettori all’interno
Come altri GPCR, il complesso PKD1–PKD2 deve essere spento e riciclato dopo l’attivazione. Gli autori hanno scoperto che una chinasi chiamata GRK6, che risiede alla membrana plasmatica, si associa fortemente a PKD1 quando è presente PKD2 e aiuta a rimuovere il complesso dalla superficie cellulare. Un’altra proteina, la β-arrestina 2, viene quindi reclutata al complesso attivato e accompagna sia PKD1 sia PKD2 negli endosomi precoci all’interno della cellula. Bloccare l’attività di GRK6 o la funzione della β-arrestina riduceva questa internalizzazione e ripristinava parzialmente la segnalazione delle proteine G. Questi passaggi di desensibilizzazione suggeriscono che il recettore PKD1–PKD2 è strettamente regolato, prevenendo una soppressione incontrollata del cAMP in condizioni normali.
Cosa significa per la malattia renale e oltre
Complessivamente, i risultati stabiliscono il complesso PKD1–PKD2 come una nuova classe di recettore accoppiato a proteine G—un recettore non convenzionale a 11 transmembrane che dipende da una subunità partner per il corretto posizionamento sulla superficie cellulare. Nei reni sani, i segnali WNT che agiscono attraverso questo recettore attivano proteine G inibitorie e mantengono i livelli di cAMP in un intervallo sicuro, contribuendo a conservare il diametro normale dei tubuli e a prevenire la formazione di cisti. Quando PKD1 o PKD2 sono mutati, questa via WNT–GPCR fallisce, il cAMP aumenta e le cisti si espandono. Chiarendo questo passo iniziale nel processo patologico, lo studio apre la strada a terapie mirate a ripristinare o mimare la segnalazione PKD1–PKD2, potenzialmente offrendo alternative più precise ai farmaci attuali che attenuano il cAMP in modo più ampio.
Citazione: Hardy, E.P., Haider, A.N., Patel, M.M. et al. A heteromeric TRP channel that functions as a WNT-activated G protein-coupled receptor. Nat Commun 17, 3233 (2026). https://doi.org/10.1038/s41467-026-69932-w
Parole chiave: malattia renale policistica, PKD1 PKD2, recettore accoppiato a proteine G, segnalazione WNT, cAMP