Clear Sky Science · ja
WNTで活性化されるGタンパク質共役受容体として働くヘテロマー性TRPチャネル
なぜ腎臓のタンパク質が誰にとっても重要なのか
常染色体優性多発性嚢胞腎(ADPKD)は、最も一般的な遺伝性腎疾患の一つであり、世界的に腎不全の主要な原因です。数十年にわたり、主要な遺伝子—PKD1とPKD2—は知られていましたが、それらのタンパク質が細胞内で実際に何をしているかは不明でした。本論文は、これらのタンパク質が細胞表面において異例の受容体を形成し、細胞内の古典的なシグナルスイッチに直接働きかけることを明らかにします。このシステムが通常どのようにして重要なメッセンジャー分子を制御しているかを説明することで、ADPKDで嚢胞がどのように生じるかのより明確な像を示し、疾患治療の新たな手がかりを提示します。
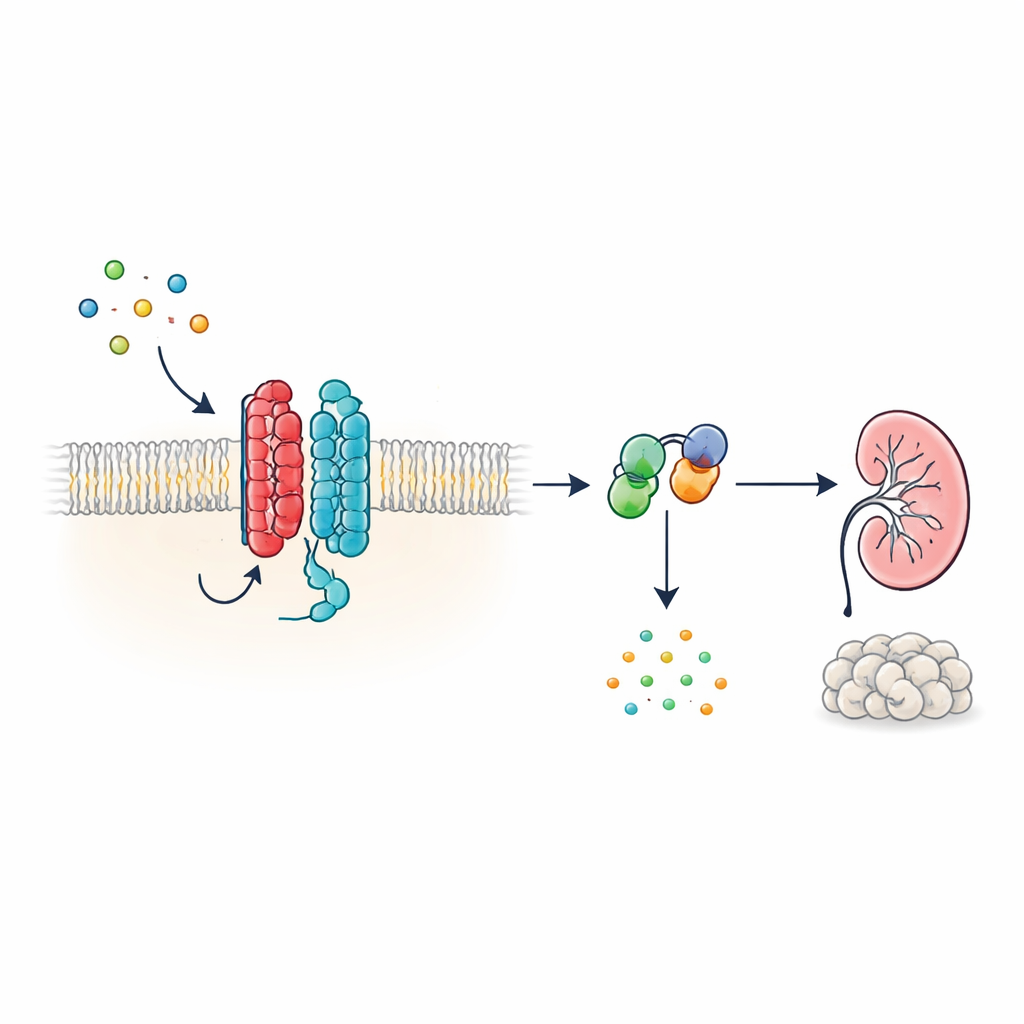
新しいタイプの細胞アンテナ
多くのホルモンや体内シグナルは、膜を七回貫通する特徴的な構造をもつ巨大な細胞表面タンパク質群であるGタンパク質共役受容体(GPCR)を介して作用します。しかしPKD1は外見が異なり、11回膜貫通を持つため、古典的受容体というよりイオンチャネルに分類されてきました。著者らは、この型破りな形状にもかかわらず、PKD1はPKD2と対をなすと真正のGPCRとして振る舞うことを示します。両者は膜上で複合体を形成し、発生や組織維持に関与する多機能なシグナル分子であるWNTタンパク質に応答します。WNTがPKD1に結合すると、この複合体は細胞内のGタンパク質を活性化し、細胞増殖や液体分泌に強く影響する小分子である環状AMP(cAMP)の濃度を変化させます。
WNTシグナルがPKD1–PKD2複合体を通じて伝わる仕組み
このシグナル伝達連鎖を追跡するため、研究者らは生細胞でGタンパク質の再配列を報告する光学的バイオセンサーを用いました。PKD1を発現するよう改変したヒト腎臓由来細胞では、いくつかのWNTファミリーメンバーが特定のGタンパク質サブユニットをパートナーから切り離す、すなわち活性化の指標であるサブユニット分離を引き起こしました。応答は阻害的なGタンパク質サブタイプであるGαi3で最も強く、関連するGαi1、Gαi2およびGαqサブユニットも関与しましたが、通常cAMPを上昇させるGタンパク質は関与しませんでした。追加のバイオセンサーは、WNT刺激がGタンパク質をPKD1に近づけるだけでなく、Gαサブユニット上でGDPがGTPに置換される化学ステップを駆動し、活性化サイクルを完了させることを示しました。これらの効果は他の既知のWNT受容体が抑制されても起き、従来のWNT受容体ではなくPKD1自体が責任を負っていることを示しています。
必須のパートナーであり門番としてのPKD2
PKD1はWNTリガンドやGタンパク質に結合できますが、それだけでは機能しません。研究チームはゲノム編集でPKD2を細胞から除去すると、WNTがもはやPKD1を介してGタンパク質活性化を駆動できなくなることを発見しました。正常なPKD2を再導入するとシグナルは回復しましたが、複合体の組立てや膜局在を乱す疾患関連のPKD2変異は回復させませんでした。顕微鏡観察とエネルギー移動測定は、PKD2がシャペロンとして働き、PKD1を細胞表面へ連れて行き、そこで安定化させることを示しました。重要なことに、PKD2のイオンチャネル活性を変える変異であってもPKD1との結合に影響しないものはGタンパク質シグナルを維持し、化学的阻害剤でイオン流を遮断してもほとんど影響がありませんでした。これは、この文脈ではPKD1–PKD2複合体が作動するイオンチャネルというより主に受容体として機能していることを示唆します。
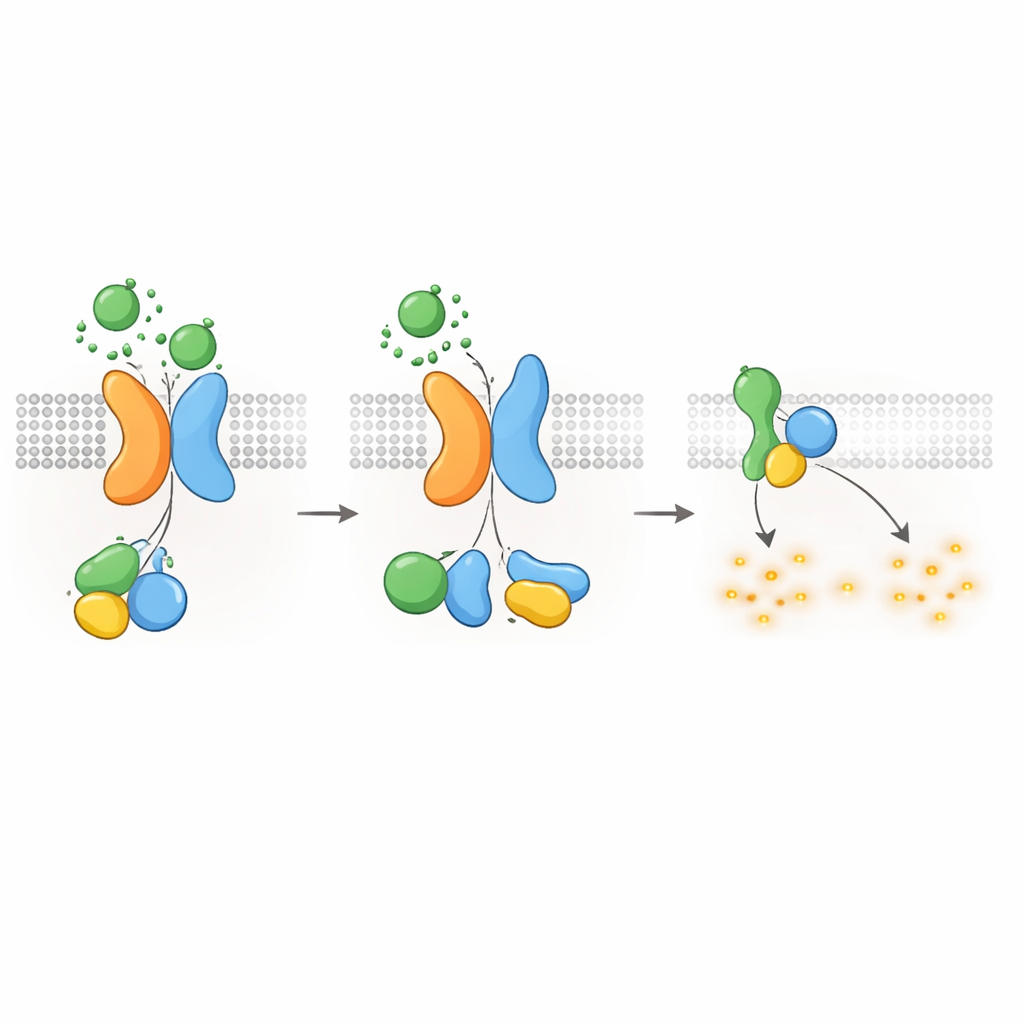
cAMP――そして嚢胞増殖――を抑える仕組み
Gαi型Gタンパク質がcAMP産生を抑えることは知られているため、著者らは次にPKD1–PKD2受容体がcAMPレベルを直接低下させ得るかを調べました。リアルタイムでcAMPを追跡する発光レポーターを用い、PKD1を発現する細胞でのWNT刺激がcAMP産生を高める化合物フォルスコリンによるcAMP上昇を遅らせることを示しました。WNTは安静時のcAMPレベルも低下させ、これら両効果は特異的にGαiタンパク質を無力化する百日咳毒素で阻害されました。WNT結合、自己プロテオリシス、またはGタンパク質相互作用を阻害する疾患原因性PKD1変異は、このcAMP低下応答を著しく弱めました。PKD2を欠く細胞もWNT依存のcAMP抑制を示さず、正常なPKD2の再導入でのみ回復しました。これらの発見は、PKD1またはPKD2の分子欠陥がcAMP抑制の喪失に直接つながり、これはADPKDにおける嚢胞成長の中心的推進因子であることを結びつけます。
シグナルの遮断と受容体の細胞内移行
他のGPCRと同様に、PKD1–PKD2複合体は活性化後にオフにされ再利用される必要があります。著者らは、細胞膜に存在するキナーゼGRK6がPKD2が存在する場合に強くPKD1と結合し、複合体を細胞表面から除去するのを助けることを見出しました。続いてβ-アレスチン2という別のタンパク質が活性化複合体にリクルートされ、PKD1とPKD2の両方を細胞内部の初期エンドソームへ連れて行きます。GRK6活性やβ-アレスチン機能を阻害するとこの内在化が減少し、Gタンパク質シグナルが部分的に回復しました。これらの脱感作ステップは、PKD1–PKD2受容体が厳密に調節されており、正常条件下でcAMPの過度な抑制が起きるのを防いでいることを示唆します。
腎疾患とその周辺領域への意義
総じて、本研究はPKD1–PKD2複合体を新しいクラスのGタンパク質共役受容体として確立します—細胞表面での適切な配置にパートナーサブユニットを必要とする非定型の11膜貫通受容体です。健常な腎臓では、この受容体を介するWNTシグナルが阻害性Gタンパク質を活性化し、cAMPレベルを安全な範囲に保つことで、尿細管の直径を維持し嚢胞形成を防ぎます。PKD1またはPKD2が変異すると、このWNT–GPCR経路が働かなくなり、cAMPが上昇して嚢胞が拡大します。この疾患過程の初期段階を明らかにすることで、本研究はPKD1–PKD2シグナルを回復または模倣することを目指した治療法への道を開き、cAMPをより広く抑える現在の薬剤に比べてより精密な代替を提供する可能性があります。
引用: Hardy, E.P., Haider, A.N., Patel, M.M. et al. A heteromeric TRP channel that functions as a WNT-activated G protein-coupled receptor. Nat Commun 17, 3233 (2026). https://doi.org/10.1038/s41467-026-69932-w
キーワード: 多発性嚢胞腎, PKD1 PKD2, Gタンパク質共役受容体, WNTシグナル, cAMP