Clear Sky Science · fr
Un canal TRP hétéromérique qui fonctionne comme un récepteur couplé aux protéines G activé par WNT
Pourquoi une protéine rénale concerne tout le monde
La maladie polykystique autosomique dominante (MPAD) est l’un des troubles rénaux héréditaires les plus fréquents et une cause majeure d’insuffisance rénale dans le monde. Pendant des décennies, les chercheurs ont identifié les gènes principaux impliqués — PKD1 et PKD2 — sans comprendre précisément la fonction de leurs produits protéiques à l’intérieur des cellules. Cet article montre que ces protéines forment à la surface cellulaire un récepteur inhabituel qui communique directement avec les commutateurs de signalisation classiques à l’intérieur de la cellule. En expliquant comment ce système contribue normalement à réguler un messager clé, l’étude éclaire la façon dont se forment les kystes dans la MPAD et ouvre des pistes pour de nouvelles approches thérapeutiques.
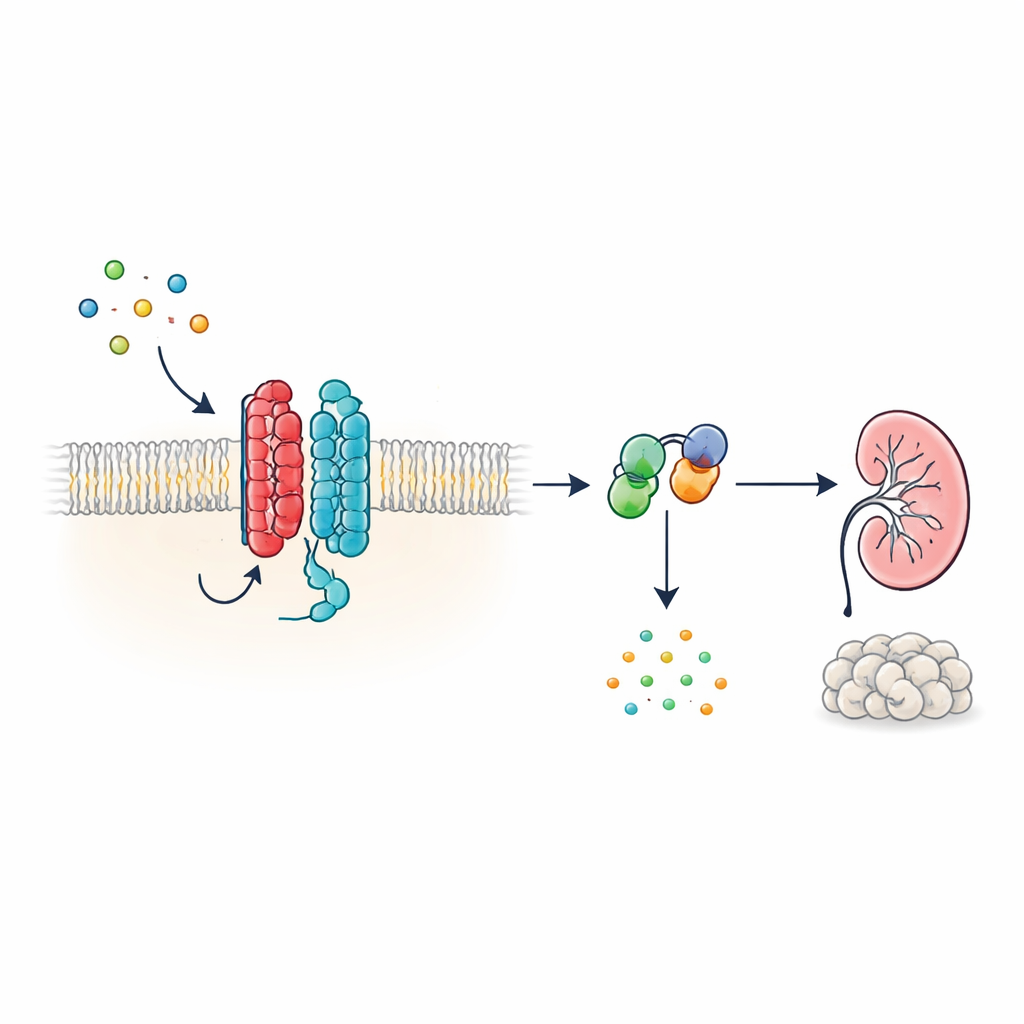
Un nouvel type d’antenne cellulaire
Beaucoup d’hormones et de signaux dans l’organisme agissent via des récepteurs couplés aux protéines G (RCPG), une vaste famille de protéines membranaires caractérisées par une structure en sept segments traversant la membrane. PKD1 diffère toutefois : elle présente 11 segments et a été classée avec les canaux ioniques plutôt qu’avec les récepteurs classiques. Les auteurs montrent que, malgré cette architecture non conventionnelle, PKD1 se comporte comme un véritable RCPG lorsqu’elle est associée à PKD2. Ensemble, ces deux protéines forment un complexe membranaire qui répond aux protéines WNT — des molécules de signalisation polyvalentes impliquées dans le développement et le maintien des tissus. Lorsque les WNT se lient à PKD1, le complexe active des protéines G intracellulaires et modifie les niveaux d’AMP cyclique (AMPc), une petite molécule qui influence fortement la croissance cellulaire et la sécrétion de liquide.
Comment les signaux WNT transitent par le complexe PKD1–PKD2
Pour tracer cette chaîne de signalisation, les chercheurs ont utilisé des biosenseurs optiques qui rapportent la réorganisation des protéines G dans des cellules vivantes. Dans des cellules d’origine rénale humaine modifiées pour exprimer PKD1, plusieurs membres de la famille WNT ont provoqué la dissociation de sous-unités spécifiques des protéines G — un signe caractéristique d’activation. La réponse était la plus forte pour un sous-type inhibiteur de protéine G, connu sous le nom de Gαi3, et impliquait aussi les sous-unités apparentées Gαi1, Gαi2 et Gαq, mais pas les protéines G qui augmentent habituellement l’AMPc. D’autres biosenseurs ont montré que la stimulation par WNT rapprochait non seulement les protéines G de PKD1, mais favorisait aussi l’étape chimique où le GDP est échangé contre le GTP sur la sous-unité Gα, complétant ainsi le cycle d’activation. Ces effets se produisaient même lorsque d’autres récepteurs WNT connus étaient supprimés, ce qui indique que c’est PKD1 lui‑même, plutôt que les récepteurs WNT classiques, qui était responsable.
PKD2, partenaire essentiel et gardien
Même si PKD1 peut lier à la fois les ligands WNT et les protéines G, elle n’agit pas seule. L’équipe a utilisé l’édition du génome pour supprimer PKD2 des cellules et a constaté que WNT ne pouvait plus activer les protéines G via PKD1. La réintroduction d’un PKD2 normal a restauré la signalisation, mais des variantes de PKD2 associées à la maladie, qui perturbent l’assemblage du complexe ou son adressage à la membrane, n’ont pas réussi à le faire. La microscopie et des mesures de transfert d’énergie ont montré que PKD2 joue le rôle d’un chaperon, escortant PKD1 vers la surface cellulaire et la stabilisant là. Fait important, des mutations qui modifient l’activité de canal ionique de PKD2 sans perturber son association avec PKD1 n’affectaient pas la signalisation via les protéines G, et bloquer le flux ionique avec un inhibiteur chimique avait peu d’effet. Cela indique que, dans ce contexte, le complexe PKD1–PKD2 fonctionne principalement comme un récepteur plutôt que comme un canal ionique actif.
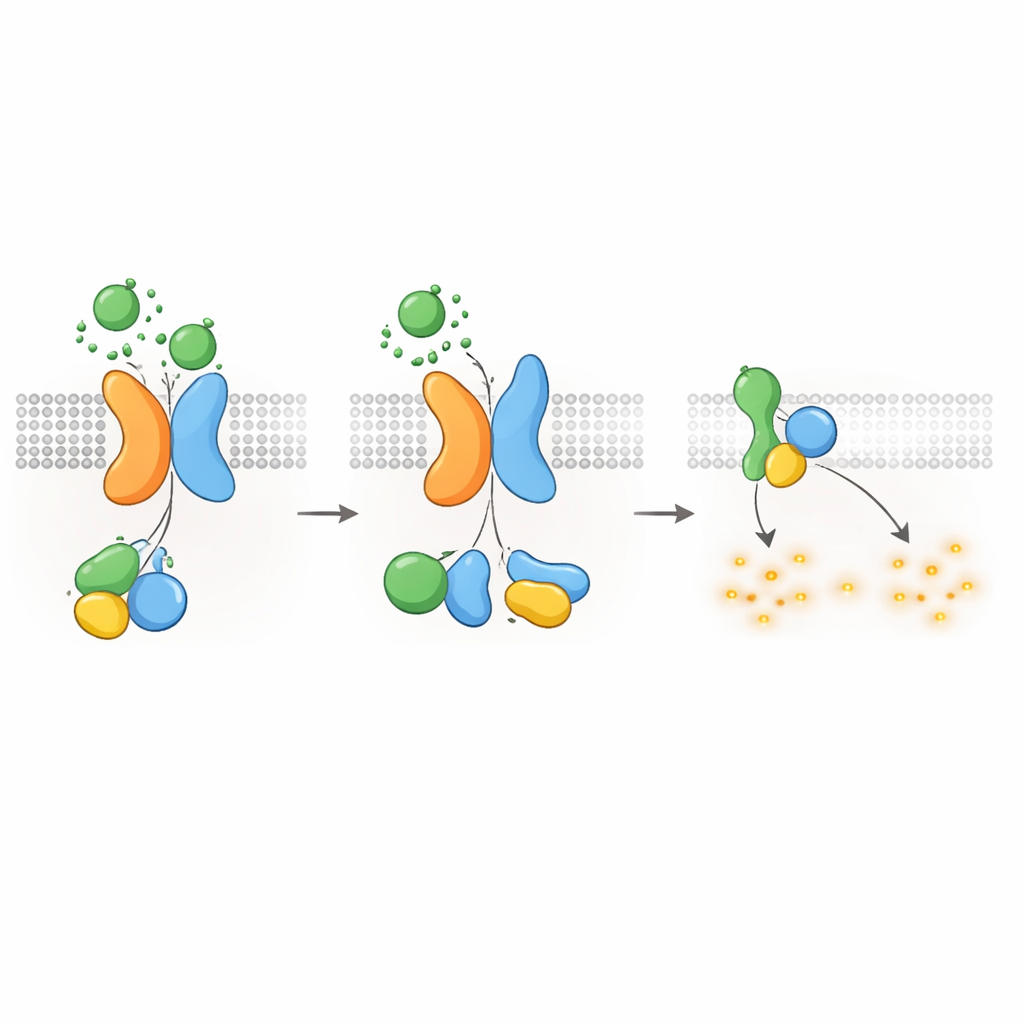
Maintenir l’AMPc — et la croissance des kystes — sous contrôle
Étant donné que les protéines G de type Gαi sont connues pour réduire la production d’AMPc, les auteurs ont ensuite vérifié si le récepteur PKD1–PKD2 pouvait abaisser directement les niveaux d’AMPc. À l’aide d’un traceur luminescent qui suit l’AMPc en temps réel, ils ont montré que la stimulation par WNT dans des cellules exprimant PKD1 ralentissait l’augmentation d’AMPc déclenchée par la forskoline, un composé qui stimule la production d’AMPc. Les WNT diminuaient aussi les niveaux d’AMPc au repos, et ces deux effets étaient bloqués par la toxine pertussique, qui désactive spécifiquement les protéines Gαi. Des mutations pathogènes de PKD1 — affectant la liaison aux WNT, le clivage autoprotéolytique ou l’interaction avec les protéines G — affaiblissaient fortement cette réponse d’abaissement de l’AMPc. Les cellules dépourvues de PKD2 n’affichaient pas non plus la suppression de l’AMPc dépendante des WNT, sauf si un PKD2 normal était réintroduit. Ces résultats relient directement les défauts moléculaires de PKD1 ou PKD2 à la perte de contrôle de l’AMPc, un moteur central de l’élargissement des kystes dans la MPAD.
Éteindre le signal et internaliser les récepteurs
Comme pour d’autres RCPG, le complexe PKD1–PKD2 doit être éteint et recyclé après activation. Les auteurs ont découvert qu’une kinase appelée GRK6, située à la membrane plasmique, s’associe fortement à PKD1 lorsque PKD2 est présente et aide à retirer le complexe de la surface cellulaire. Une autre protéine, la β-arrestine 2, est alors recrutée au complexe activé et conduit PKD1 et PKD2 vers des endosomes précoces à l’intérieur de la cellule. Bloquer l’activité de GRK6 ou la fonction de la β-arrestine réduisait cette internalisation et restaurait partiellement la signalisation via les protéines G. Ces étapes de désensibilisation suggèrent que le récepteur PKD1–PKD2 est étroitement régulé, empêchant une suppression excessive de l’AMPc dans des conditions normales.
Ce que cela signifie pour la maladie rénale et au-delà
Ensemble, les résultats établissent le complexe PKD1–PKD2 comme une nouvelle classe de récepteur couplé aux protéines G — un récepteur non conventionnel à 11 domaines transmembranaires qui dépend d’une sous‑unité partenaire pour un positionnement correct à la surface cellulaire. Dans des reins sains, les signaux WNT agissant via ce récepteur activent des protéines G inhibitrices et maintiennent les niveaux d’AMPc dans une fourchette sûre, aidant à préserver le diamètre normal des tubules et à prévenir la formation de kystes. Lorsque PKD1 ou PKD2 est muté, cette voie WNT–RCPG faiblit, l’AMPc augmente et les kystes se développent. En clarifiant cette étape précoce du processus pathologique, l’étude ouvre la voie à des thérapies visant à restaurer ou à mimer la signalisation PKD1–PKD2, offrant potentiellement des alternatives plus ciblées aux médicaments actuels qui atténuent l’AMPc de façon plus générale.
Citation: Hardy, E.P., Haider, A.N., Patel, M.M. et al. A heteromeric TRP channel that functions as a WNT-activated G protein-coupled receptor. Nat Commun 17, 3233 (2026). https://doi.org/10.1038/s41467-026-69932-w
Mots-clés: maladie polykystique des reins, PKD1 PKD2, récepteur couplé aux protéines G, signalisation WNT, AMPc