Clear Sky Science · ru
Снятие репрессии фосфатазы белка 5 организатором шаперонов HOP1 активирует иммунитет NLR у растений
Как растения выбирают между ростом и самозащитой
Растения постоянно сталкиваются с потенциальными захватчиками — бактериями, грибами и вирусами — но они не могут просто прекратить рост, чтобы сражаться с каждой угрозой. В этой статье раскрывается, как крошечная молекулярная система «тормоза и спускового крючка» в модельном растении Arabidopsis помогает решить, когда запустить экстремальную иммунную реакцию, которая может спасти растение, но также подавить его рост. Понимание этого переключателя даёт представление о том, как можно селекционно или с помощью инженерии сделать сельскохозяйственные культуры устойчивыми к болезням, не платя слишком высокую цену в виде снижения урожая.
Повседневные тревоги против аварийных сирен
У растений есть две основные системы охраны. Первая — быстрая, общая сигнализация, которая обнаруживает характерные признаки микробов на поверхности клетки и включает защитные ответы. Вторая, более радикальная ступень опирается на внутренние сторожевые молекулы, называемые рецепторами NLR, которые могут запустить мощную, часто смертельную, защиту в клетках, где обнаружены патогены. В здоровом Arabidopsis цепочка сигналов из трёх ферментов — в совокупности называемая каскадом MEKK1–MKK1/2–MPK4 — одновременно передаёт первую тревогу и тонко удерживает один особый рецептор NLR, называемый SUMM2, в неактивном состоянии. Когда эта цепочка нарушается мутацией или бактериальным эффекторным белком, у растений возникают спонтанная гибель клеток, высокая активность генов защиты и сильный карликовый рост — всё это признаки неконтролируемого иммунитета.
Пробуждение скрытой ферменты
Авторы обнаружили, что фосфатаза, называемая LET7, растительная версия консервативной белковой PP5, находится в центре этого неконтролируемого ответа. Сама по себе LET7 устроена как сложный карманный нож: одна область (TPR-домен) загибается над каталитической областью и сильно подавляет её активность. Генетические скрининги показали, что растения, лишённые LET7, больше не испытывают летальных симптомов, возникающих при молчании верхней сигнальной цепочки, что помещает LET7 ниже по потоку этой каскадной цепи. Биохимические тесты выявили, что когда каталитическая область LET7 освобождается или становится гиперактивной из‑за определённых мутаций, растения развивают сильные защитные ответы и гибель клеток, тогда как каталитически неактивная версия не может инициировать эти эффекты. Таким образом, фосфатазная активность LET7 необходима и достаточна для запуска автоиммунитета.
Молекулярный организатор отпускaет тормоз
Как же открывается «карманный нож»? Группа идентифицировала ко-шаперонный белок HOP1 как ключевого организатора. HOP1 обычно помогает белкам шока тепла сворачиваться и стабилизировать другие белки. Здесь он прикрепляется к TPR‑региону LET7 и конкурирует с внутренним контактом, который удерживает LET7 в закрытом состоянии. Структурное моделирование и точечные замены аминокислот в обоих партнёрах показали, что эта поверхность связывания тонко настроена: ослабление контакта между HOP1 и LET7 снижает активацию LET7. Когда каскад MEKK1–MKK1/2–MPK4 цел, ассоциация между HOP1 и LET7 слаба и LET7 остаётся репрессированной. Когда каскад нарушается — из‑за мутаций, избыточного количества другого каркасного белка или бактериального эффектора — взаимодействие HOP1–LET7 усиливается, освобождая каталитическую активность LET7. Примечательно, что человеческие аналоги HOP1 и PP5 способны заместить растительную пару, что указывает на сохранённый механизм активации между царствами живого.
Перекусывая ключевой переключатель и охраняя датчик
После активации LET7 нацелен на киназу CRCK3, фермент, ранее связанный с тем же иммунным путём. CRCK3 существует в фосфорилированной и дефосфорилированной формах, которые можно различить как две полосы на гелях белков. В растительных клетках и в тестах в пробирке активная LET7 выборочно удаляет фосфатные метки с CRCK3, тогда как неактивная LET7 этого не делает. Усиление активности LET7 или её совместная экспрессия с CRCK3 усиливают де-фосфорилирование и приводят к гибели клеток и классическим маркёрам иммунитета, но эти эффекты ослабляются при отсутствии HOP1 или рецептора NLR SUMM2. Параллельно авторы обнаружили, что и LET7, и HOP1 через свои TPR‑регионы физически связываются с SUMM2 и повышают его уровень белка, не стабилизируя при этом широко другие рецепторы NLR. Таким образом, тот же модуль, который удаляет фосфат с CRCK3, также помогает поддерживать сенсор, считывающий этот сигнал.
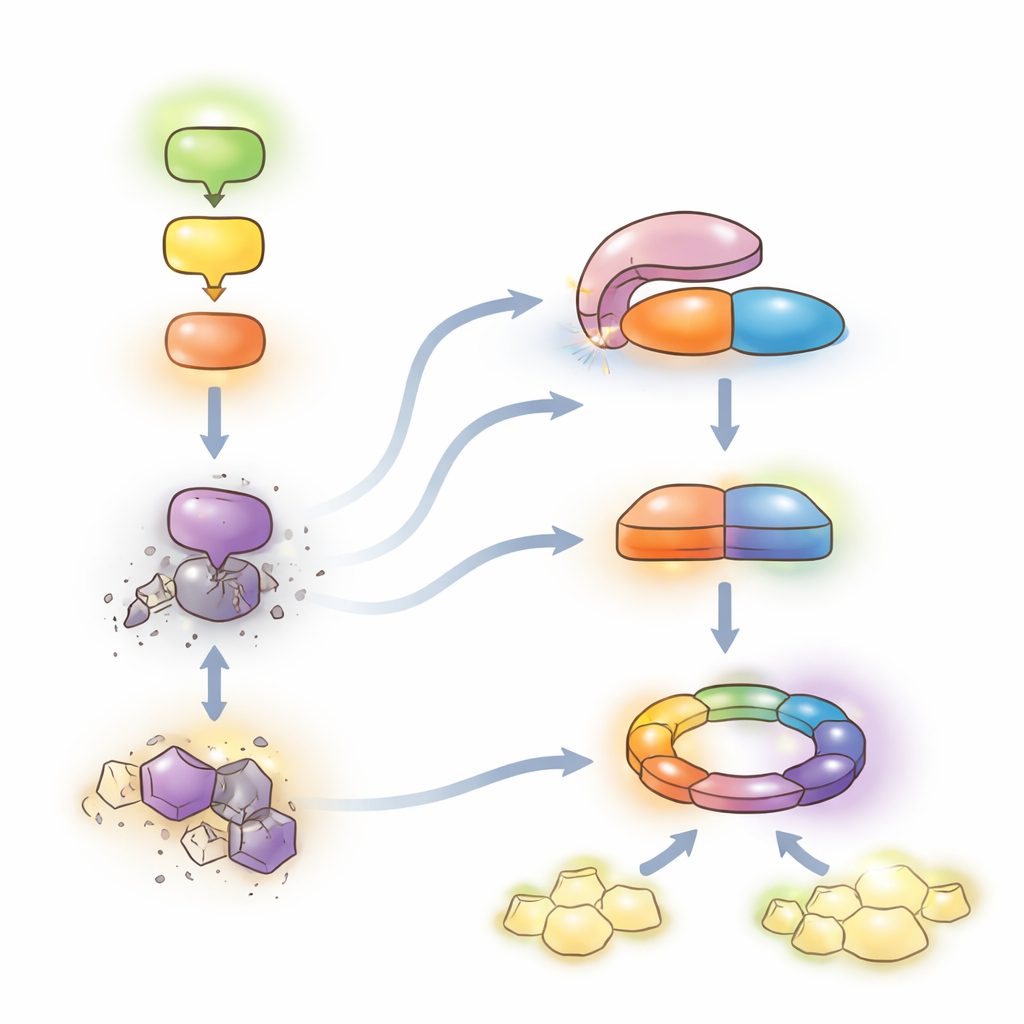
Тонкий баланс между защитой и ростом
Вместе результаты поддерживают модель, в которой каскад MEKK1–MKK1/2–MPK4 сдерживает мощную вторую линию защиты. Когда эта цепочка цела, LET7 остаётся автомодулировано неактивной, а уровни SUMM2 поддерживаются в разумных пределах, что позволяет растениям в основном полагаться на свою повседневную систему оповещения и продолжать рост. Когда патогены нарушают цепочку, HOP1 взаимодействует с LET7, разблокируя её фосфатазную активность. LET7 затем дефосфорилирует CRCK3 и, вместе с HOP1, стабилизирует SUMM2, переводя систему в режим полного NLR‑опосредованного иммунитета и локальной гибели клеток — высокозащитного, но затратного для роста. Раскрывая этот консервативный механизм «снятия репрессии» для ферментов PP5, работа описывает молекулярный рычаг, который в принципе можно регулировать в культурах для усиления устойчивости к болезням при минимизации вредного автоиммунитета.
Цитирование: Yan, Y., Zhao, Z., Yeo, IC. et al. De-repression of protein phosphatase 5 by the chaperone organizer HOP1 activates plant NLR immunity. Nat. Plants 12, 802–817 (2026). https://doi.org/10.1038/s41477-026-02253-4
Ключевые слова: иммунитет растений, фосфатаза белка 5, рецепторы NLR, сигнальная трансдукция, Arabidopsis