Clear Sky Science · ja
シャペロンオーガナイザーHOP1によるタンパク質ホスファターゼ5の脱抑制が植物NLR免疫を活性化する
成長と自己防御を植物はどう選ぶか
植物は常に細菌、真菌、ウイルスといった侵入者に晒されていますが、すべての脅威に対して成長を放棄して戦うわけにはいきません。本論文はモデル植物シロイヌナズナにおいて、小さな分子レベルの「ブレーキと作動装置」システムが、植物を救う一方で成長を阻害しうる極端な免疫反応をいつ発動するかを決める仕組みを明らかにします。このスイッチの理解は、収量への負担を抑えつつ作物の病害抵抗性を高める育種や遺伝子改変の手がかりを与えます。
日常の警報と非常時のサイレン
植物は主に二つの防御システムを運用しています。第一は細胞表面で一般的な微生物の指紋を検出する迅速で広範な警報系で、防御応答を立ち上げます。第二はより劇的で、NLR受容体という内部の哨兵に依存し、病原体が検出された細胞で強力かつしばしば致死的な防御を誘導します。健常なシロイヌナズナでは、MEKK1–MKK1/2–MPK4と総称される三段の酵素連鎖が第一の警報を伝えると同時に、特定のNLR受容体であるSUMM2が不意に作動するのを静かに抑えています。この連鎖が変異や細菌エフェクター蛋白質によって破壊されると、自然発生的な細胞死、防御遺伝子の高発現、重度の矮小化といった暴走した免疫の兆候が現れます。
埋もれていた酵素の覚醒
著者らは、LET7と呼ばれるホスファターゼ(保存的なタンパク質PP5の植物相同体)がこの暴走応答の中心にあることを発見しました。LET7自体は折りたたまれたポケットナイフのような構造で、ある領域(TPRドメイン)が触媒領域に折り返して作用を大きく抑えています。遺伝学的スクリーニングにより、LET7を欠く植物は上流のシグナル伝達が沈黙した際に生じる致死的症状を示さなくなり、LET7がそのカスケードの下流に位置することが示されました。生化学的検査では、LET7の触媒領域が解放されるか特定の変異で過活性化されると植物は強い防御応答と細胞死を示す一方、触媒不活化型はこれらを引き起こせないことが明らかになりました。したがって、LET7のホスファターゼ活性は自己免疫を駆動するために必要かつ十分です。
分子オーガナイザーがブレーキを解く
ではそのポケットナイフはどう開くのか。研究チームは共シャペロン(コシャペロン)タンパク質HOP1を重要なオーガナイザーとして同定しました。HOP1は通常ヒートショックタンパク質の働きを助け、他のタンパク質の折り畳みと安定化に寄与します。本系ではHOP1がLET7のTPR領域に取り付き、LET7を閉じた状態に保つ内部の接触を競合的に乗り越えます。構造モデリングと両者に対する厳密なアミノ酸置換実験は、この結合面が精密に調整されていることを示しました:HOP1とLET7間の接触を弱めるとLET7の活性化は低下します。MEKK1–MKK1/2–MPK4鎖が健全なときはHOP1とLET7の結合が低くLET7は抑制されたままです。一方その鎖が変異や別の足場タンパク質の過剰、あるいは細菌エフェクターによって乱れると、HOP1–LET7相互作用が強まりLET7の触媒活性が解放されます。驚くべきことに、ヒトのHOP1およびPP5の相同体が植物の組み合わせを代替でき、王国を超えた保存的な活性化機構が示唆されます。
重要なスイッチを切り、センサーを守る
一旦活性化したLET7は、同じ免疫経路に関連づけられていたキナーゼCRCK3を標的とします。CRCK3はリン酸化型と脱リン酸化型の二形態があり、タンパク質ゲル上で二本のバンドとして識別できます。植物細胞およびin vitroのアッセイで、活性LET7は選択的にCRCK3のリン酸基を除去し、不活性LET7はそれを行いません。LET7活性を高めるかCRCK3と共発現させると脱リン酸化が促進され、細胞死と典型的な免疫マーカーが誘導されますが、これらの効果はHOP1またはNLR受容体SUMM2が欠けていると弱まります。同時に、著者らはLET7とHOP1がそれぞれのTPR領域を介してSUMM2に物理的に結合し、他のNLR受容体を幅広く安定化することなくSUMM2のタンパク質量を増加させることを見いだしました。つまり、CRCK3からリン酸を除去する同じモジュールが、その信号を読むセンサーを維持する働きも持ち合わせているのです。
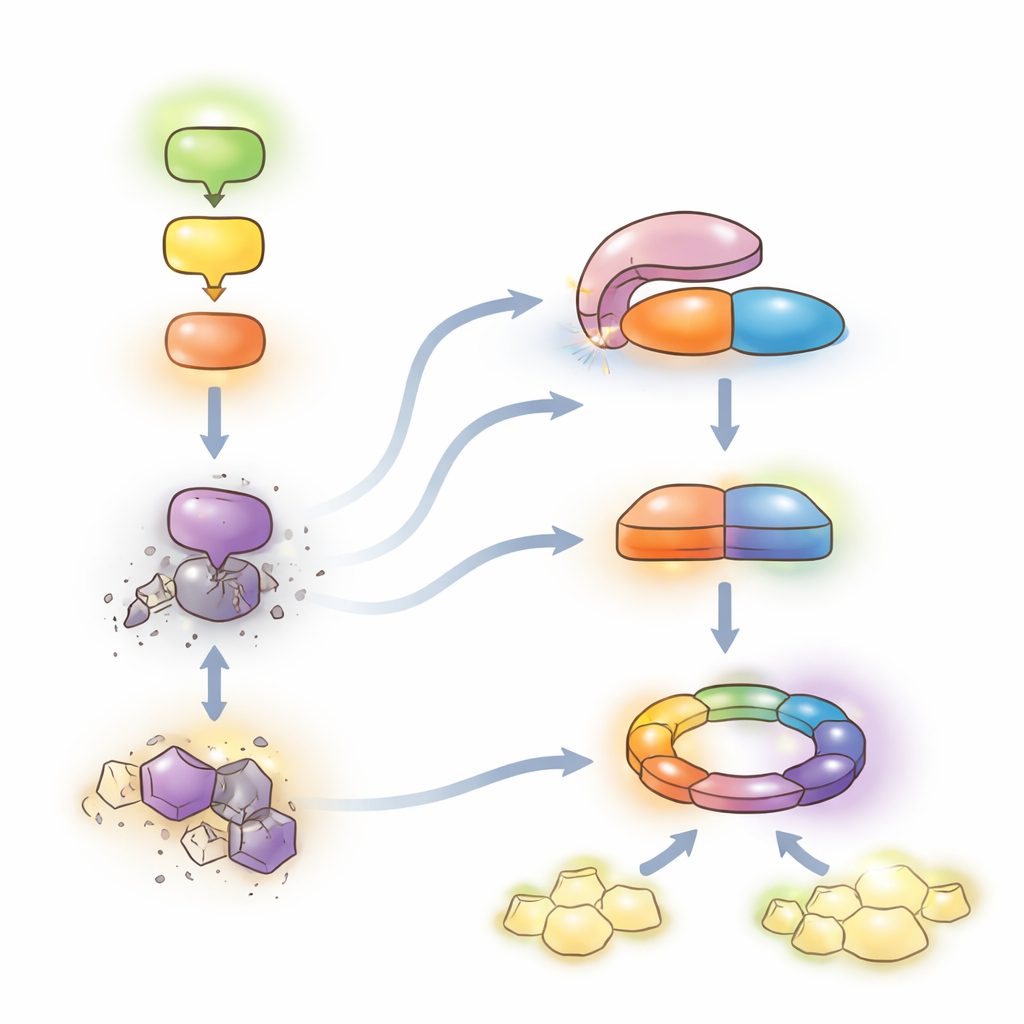
防御と成長の微妙な均衡
これらの知見は、MEKK1–MKK1/2–MPK4連鎖が強力な第二線の防御を抑制しているというモデルを支持します。鎖が健全である限り、LET7は自己抑制されSUMM2レベルは抑えられ、植物は日常の警報系に主に依存しつつ成長を続けられます。病原体がこの連鎖を乱すと、HOP1がLET7に結合してそのホスファターゼ活性を解放します。LET7はCRCK3を脱リン酸化し、HOP1とともにSUMM2を安定化させ、NLR媒介の完全な免疫と局所的な細胞死――非常に保護的だが成長には高い代償を伴う――へとシステムを傾けます。PP5酵素のこの保存された“脱抑制”機構を明らかにしたことで、作物において有害な自己免疫を最小限に抑えつつ病害抵抗性を強化するために調整可能な分子レバーが示されました。
引用: Yan, Y., Zhao, Z., Yeo, IC. et al. De-repression of protein phosphatase 5 by the chaperone organizer HOP1 activates plant NLR immunity. Nat. Plants 12, 802–817 (2026). https://doi.org/10.1038/s41477-026-02253-4
キーワード: 植物免疫, タンパク質ホスファターゼ5, NLR受容体, シグナル伝達, シロイヌナズナ