Clear Sky Science · zh
H3K14ub–H3K9me3 反馈回路调控裂殖酵母的异染色质扩展与遗传
为何这个微观开关很重要
在我们每个细胞内部,长长的 DNA 链被精心包装和调控,以确保恰当的基因在恰当的时间被开启或关闭。其中一项重要的控制机制是“异染色质”——高度致密的 DNA 区域,可将基因保持沉默并维持基因组稳定。这项在裂殖酵母中进行的研究揭示了一个隐藏的分子反馈回路,它有助于异染色质沿染色体扩展并在细胞分裂时得以维持。由于类似系统在人类中也存在,理解该回路可为细胞如何维持基因组稳定、以及其失效如何促成疾病甚至癌症提供线索。
基因组中的安静街区
并非所有 DNA 区域都同样活跃。有些区域形成异染色质,其中的基因大多被关闭。这些沉默区通过阻止重复序列异常活动、减少有害重组并塑造基因表达程序来保护基因组。异染色质具有三个关键阶段:在特定的“成核”位点启动、向邻近区域扩展、然后在细胞分裂中被传递下去。尽管启动信号已较为清楚,科学家长期困惑的是沉默状态如何可靠地扩展并持续存在,尤其是此前模型主要依赖于单一组蛋白上相对微弱的自增强标记。
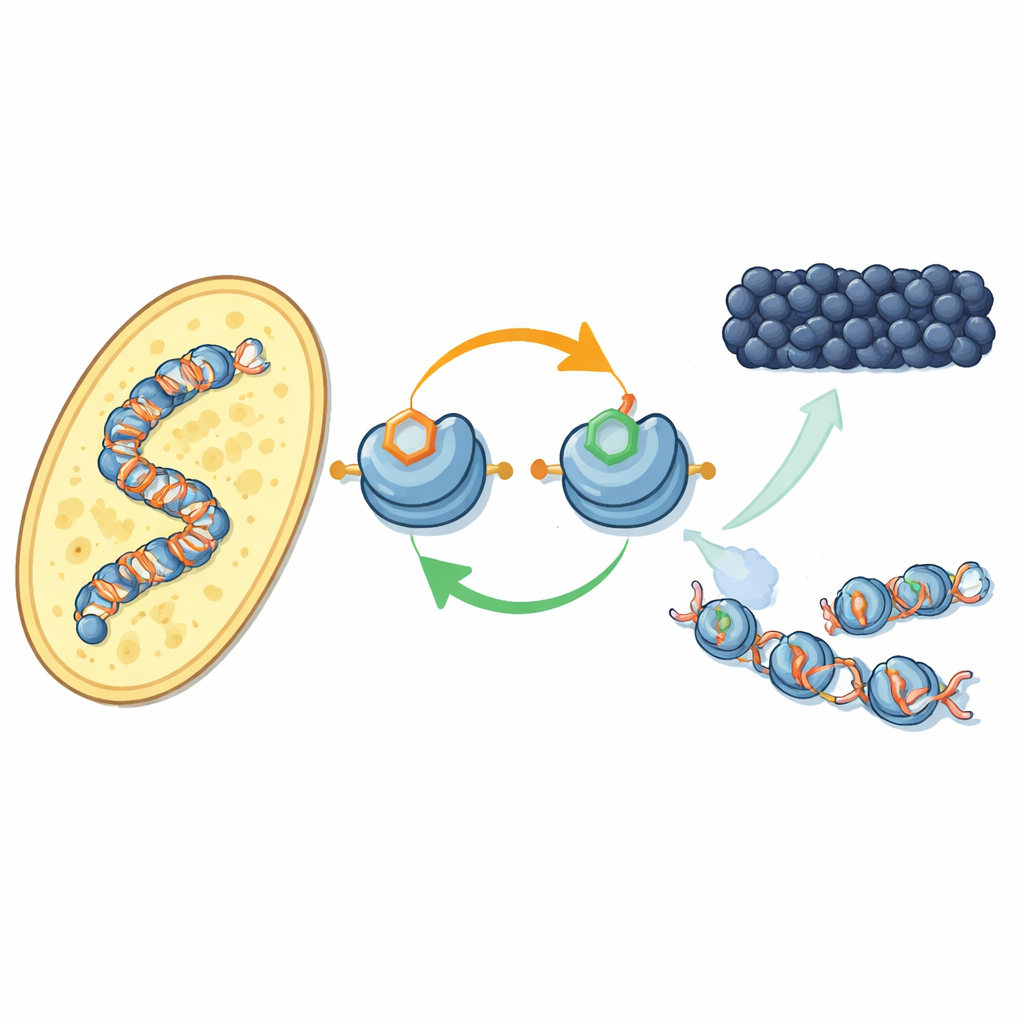
作用于 DNA 包装蛋白的新反馈回路
作者聚焦于组蛋白这一 DNA 卷绕轴上特定化学标记如何协同工作。在裂殖酵母中,一种称为 Clr4 的酶会在组蛋白 H3 的一个位置(称为 H3K9)加入甲基,这是异染色质的标志。该酶也存在于更大的复合物 CLRC 中,后者在 H3 的另一个位置(H3K14)添加一个小的泛素标签。利用纯化组分,研究者显示在 H3K14 上添加泛素能显著增强 Clr4 在完整核小体上对 H3K9 的甲基化能力,从而克服 DNA 本身的抑制作用。这一刺激效应具有高度特异性:在其他组蛋白位点的泛素化并不具有同样影响。
两种标记相互维系
转向活细胞实验,团队使用新开发的抗体绘制了 H3K14 泛素化标签在基因组中的分布。该标记几乎与 H3K9 甲基化重叠——出现在着丝粒、端粒和一个沉默的交配类型区等经典异染色质区域。当他们禁用甲基标记或放置该标记的酶时,泛素标记也随之消失,反之亦然。两种标记都需要将 CLRC 复合物锚定在染色质上。综合来看,这揭示了一个正反馈回路:泛素化促进甲基化,而这对标记又稳定负责放置它们的复合物,帮助沉默状态在邻近核小体间扩展并在 DNA 复制后恢复。
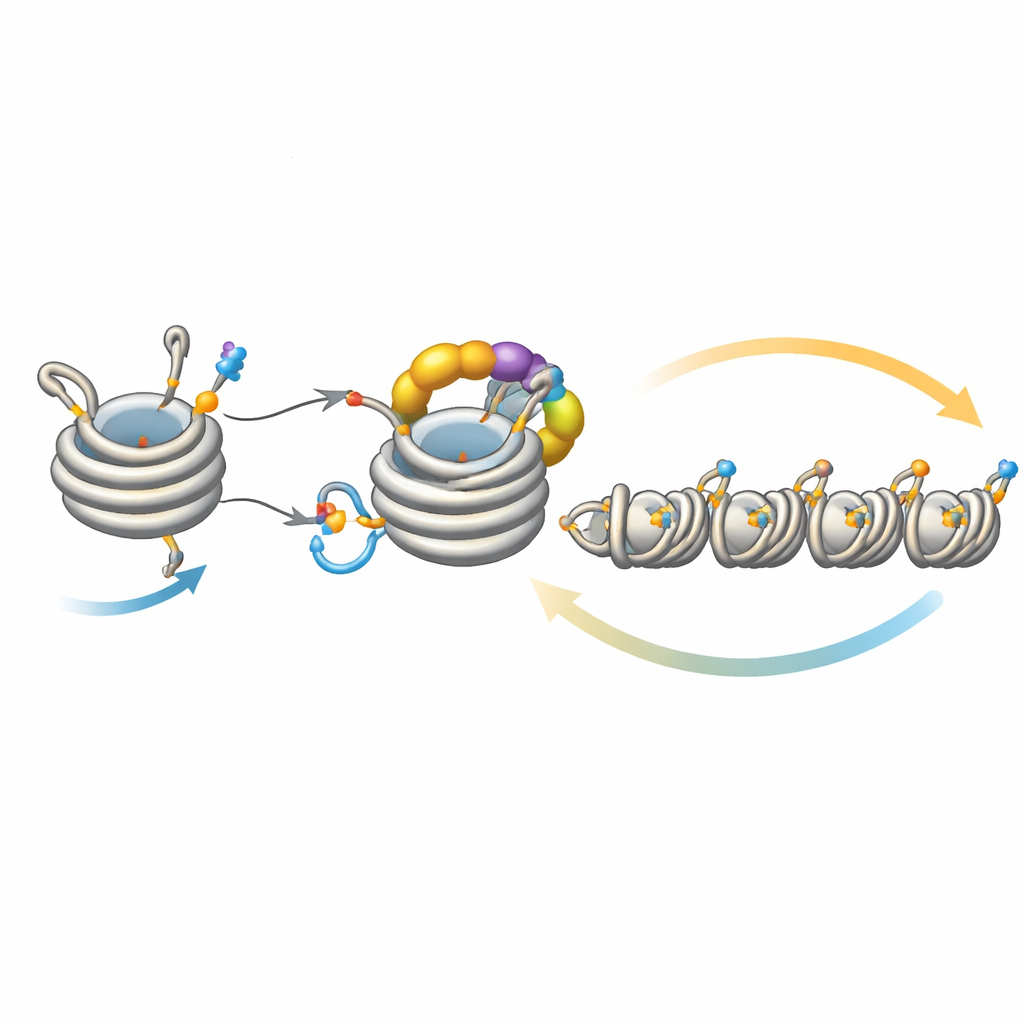
对沉默扩散的精细调控
该回路并非孤立运行。一种去乙酰化酶 Clr3 通过去除同一位点 H3K14 的乙酰基,为泛素化做准备;当这一步受损时,反馈减弱,异染色质在扩展或遗传时遇到困难,尤其是在远离成核位点的区域。相反,也有若干酶起到刹车作用。Mst2 添加阻碍泛素化的乙酰基,而 Epe1 则去除 H3K9 的甲基。移除这些刹车会导致异染色质过度扩展、沉默必需基因并使细胞承受压力。在这种压力下,细胞会触发一种适应性响应,降低 Clr4 的含量,从而抑制过度活跃的反馈回路。遗传学实验表明,强化或削弱回路不同环节可以拯救或消除扩展与遗传缺陷,凸显该系统的精细平衡。
超出酵母的意义
总体而言,研究结果表明异染色质的维持并非依赖单一微弱的“读-写”反应,而是依赖于组蛋白上泛素化、去乙酰化和甲基化的整合电路。在裂殖酵母中,该 H3K14ub–H3K9me3 反馈回路形成了一个双稳态开关:一旦某一区域跨过这些标记的阈值,它倾向于保持沉默并继续扩展,而未达到阈值的区域则不会完全进入异染色质状态。由于这两种组蛋白标记及许多相关酶在哺乳动物中是保守的,类似的逻辑可能有助于在人类细胞中维持稳定而又可调的基因沉默模式。了解这套回路最终或能为调控染色质提供新思路——例如在衰老细胞中稳定基因组,或在癌症治疗中选择性唤醒沉默区域。
引用: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
关键词: 异染色质, 组蛋白修饰, 表观遗传遗传, 染色质反馈回路, 裂殖酵母