Clear Sky Science · en
An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast
Why this microscopic switch matters
Inside every one of our cells, long strands of DNA are carefully packed and managed so that the right genes turn on or off at the right time. A major part of this control system is "heterochromatin," tightly packed DNA that keeps certain stretches of the genome silent and stable. This study, carried out in fission yeast, uncovers a hidden molecular feedback circuit that helps heterochromatin spread along chromosomes and persist when cells divide. Because similar systems exist in humans, understanding this circuit offers clues to how cells maintain genome stability and how its failure can contribute to disease and even cancer.
A quiet neighborhood in the genome
Not all DNA is equally active. Some regions form heterochromatin, where genes are largely switched off. These silent stretches protect the genome by keeping repetitive DNA from misbehaving, limiting harmful recombination, and shaping gene expression programs. Heterochromatin has three key phases: it starts at special "nucleation" sites, spreads into neighboring regions, and is then passed on through cell divisions. While the starting signals are fairly well understood, scientists have long puzzled over how the silent state reliably spreads and endures, especially given that previous models relied on a relatively weak self-reinforcing mark on one histone protein.
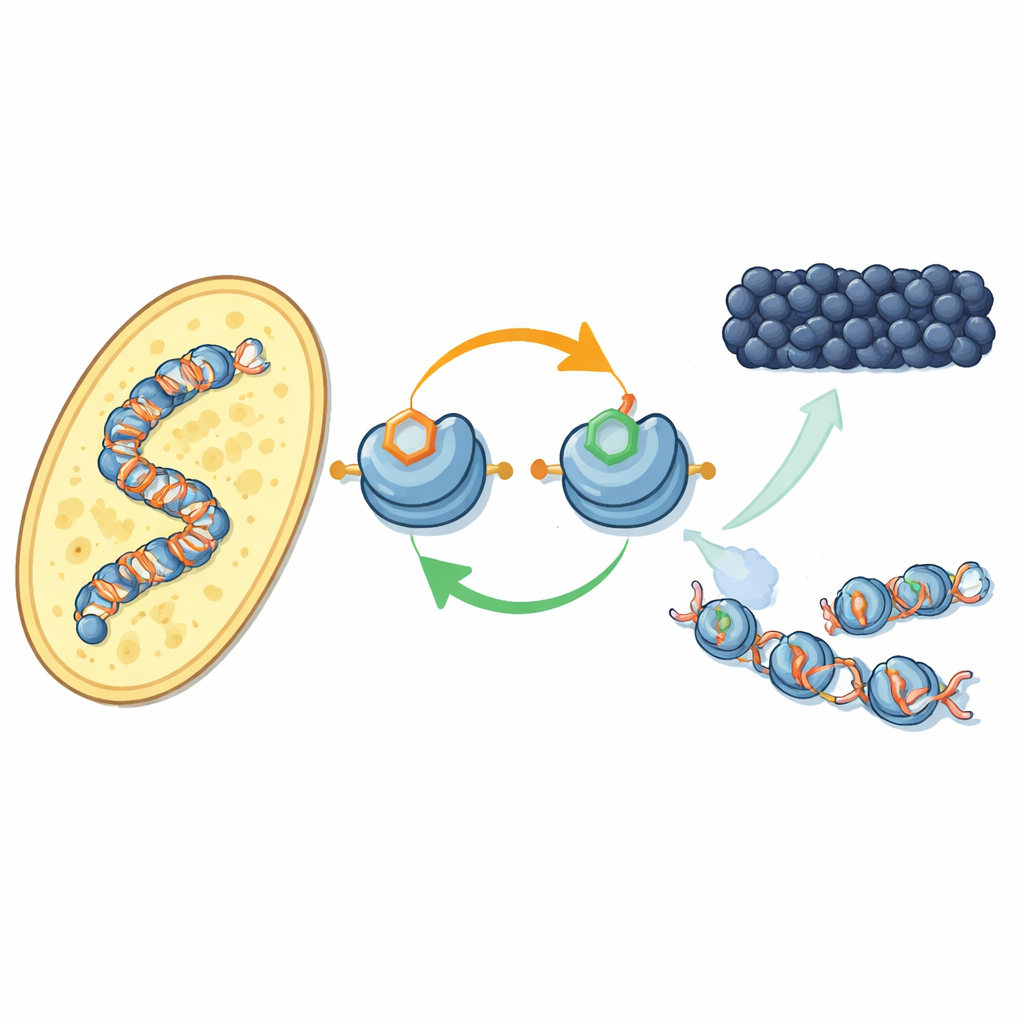
A new feedback loop on DNA packaging proteins
The authors focus on how specific chemical tags on histone proteins—the spools around which DNA is wrapped—work together. In fission yeast, an enzyme called Clr4 adds a methyl tag at one position on histone H3 (known as H3K9), a hallmark of heterochromatin. The same enzyme also sits in a larger complex, CLRC, that attaches a small ubiquitin tag at another position on H3 (H3K14). Using purified components, the researchers show that adding ubiquitin at H3K14 dramatically boosts Clr4’s ability to methylate H3K9 on full nucleosomes, overcoming an otherwise inhibitory effect of DNA. This stimulatory effect is highly specific: other ubiquitin tags on different histone positions do not have the same impact.
Two marks that sustain each other
Turning to living yeast cells, the team used a newly developed antibody to map where the H3K14 ubiquitin tag appears across the genome. It sits almost exactly where H3K9 methylation does—at centromeres, telomeres, and a silent mating-type region—classic heterochromatin territories. When they disabled either the methyl mark or the enzymes that place it, the ubiquitin mark also disappeared, and vice versa. Both tags are needed to keep the CLRC complex anchored on chromatin. Together, this reveals a positive feedback loop: the ubiquitin tag boosts methylation, and the pair of marks stabilize the very complex that places them, helping the silent state spread along neighboring nucleosomes and be restored after DNA replication.
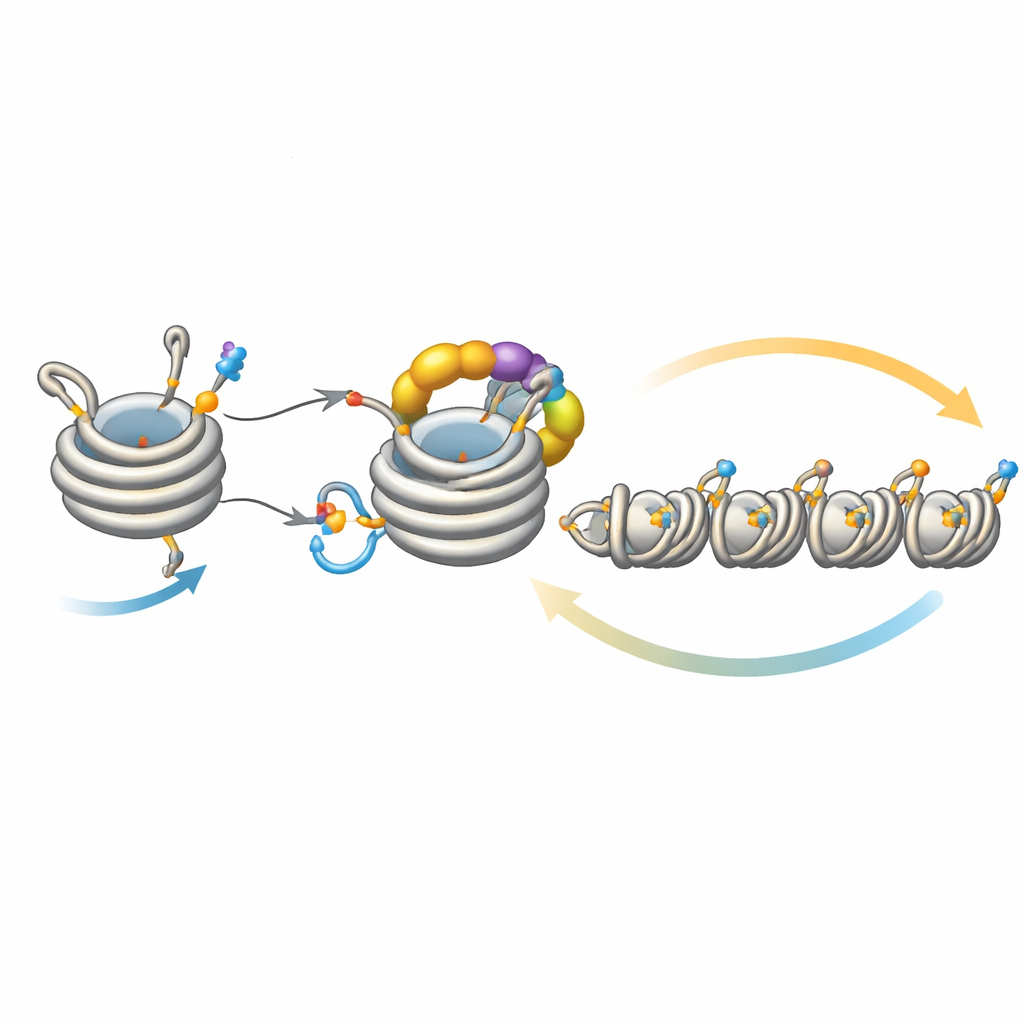
Fine-tuning the spread of silence
This loop does not operate in isolation. A deacetylase enzyme called Clr3 prepares histone H3 for ubiquitination by removing an acetyl group at the same site, H3K14; when this step is impaired, the feedback weakens and heterochromatin struggles to spread or be inherited, especially at regions far from the original nucleation sites. Conversely, several enzymes act as brakes. Mst2 adds the acetyl group that blocks ubiquitination, and Epe1 removes the methyl mark on H3K9. Removing these brakes makes heterochromatin spread too far, silencing essential genes and stressing the cells. Under such stress, cells trigger an adaptive response that reduces the amount of Clr4, dialing back the overactive feedback loop. Genetic experiments show that strengthening or weakening different arms of this circuit can rescue or abolish defects in spreading and inheritance, underscoring how delicately balanced the system is.
Implications beyond yeast cells
Together, the findings reveal that heterochromatin is maintained not by a single weak "read-write" reaction, but by an integrated circuit of ubiquitination, deacetylation, and methylation on histone proteins. In fission yeast, this H3K14ub–H3K9me3 feedback loop creates a bistable switch: once a region crosses a threshold of these marks, it tends to stay silent and spread, whereas regions that do not never fully commit to heterochromatin. Because both of these histone marks and many of the involved enzymes are conserved in mammals, a similar logic may help maintain stable yet flexible patterns of gene repression in human cells. Understanding this circuitry could eventually inform new ways to modulate chromatin—for example, to stabilize the genome in aging cells or to selectively reawaken silent regions during cancer therapy.
Citation: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
Keywords: heterochromatin, histone modification, epigenetic inheritance, chromatin feedback loop, fission yeast