Clear Sky Science · sv
Ett H3K14ub–H3K9me3-återkopplingskrets styr heterokromatinets spridning och arv i fissionsjäst
Varför denna mikroskopiska strömbrytare är viktig
Inuti varje cell paketeras och hanteras långa DNA-strängar noggrant så att rätt gener slås på eller av vid rätt tidpunkt. En huvudkomponent i detta kontrollsystem är "heterokromatin", tätt packat DNA som håller vissa delar av genomet tysta och stabila. Denna studie, utförd i fissionsjäst, avslöjar en dold molekylär återkopplingskrets som hjälper heterokromatin att sprida sig längs kromosomer och bestå när celler delar sig. Eftersom liknande system finns hos människor ger förståelsen av denna krets ledtrådar om hur celler upprätthåller genomets stabilitet och hur dess fel kan bidra till sjukdomar och även cancer.
En tyst kvarter i genomet
Inte allt DNA är lika aktivt. Vissa regioner bildar heterokromatin, där gener till stor del är avstängda. Dessa tysta partier skyddar genomet genom att hindra repetitivt DNA från att orsaka problem, begränsa skadlig rekombination och forma genuttrycksprogram. Heterokromatin har tre nyckelfaser: det börjar vid särskilda "nukleations"-platser, sprider sig till intilliggande regioner och förs sedan vidare genom celldelningar. Medan startsignalerna är rätt väl förstådda har forskare länge grubblat över hur det tysta tillståndet pålitligt sprider sig och består, särskilt eftersom tidigare modeller förlitade sig på en relativt svag självförstärkande mark på ett histonprotein.
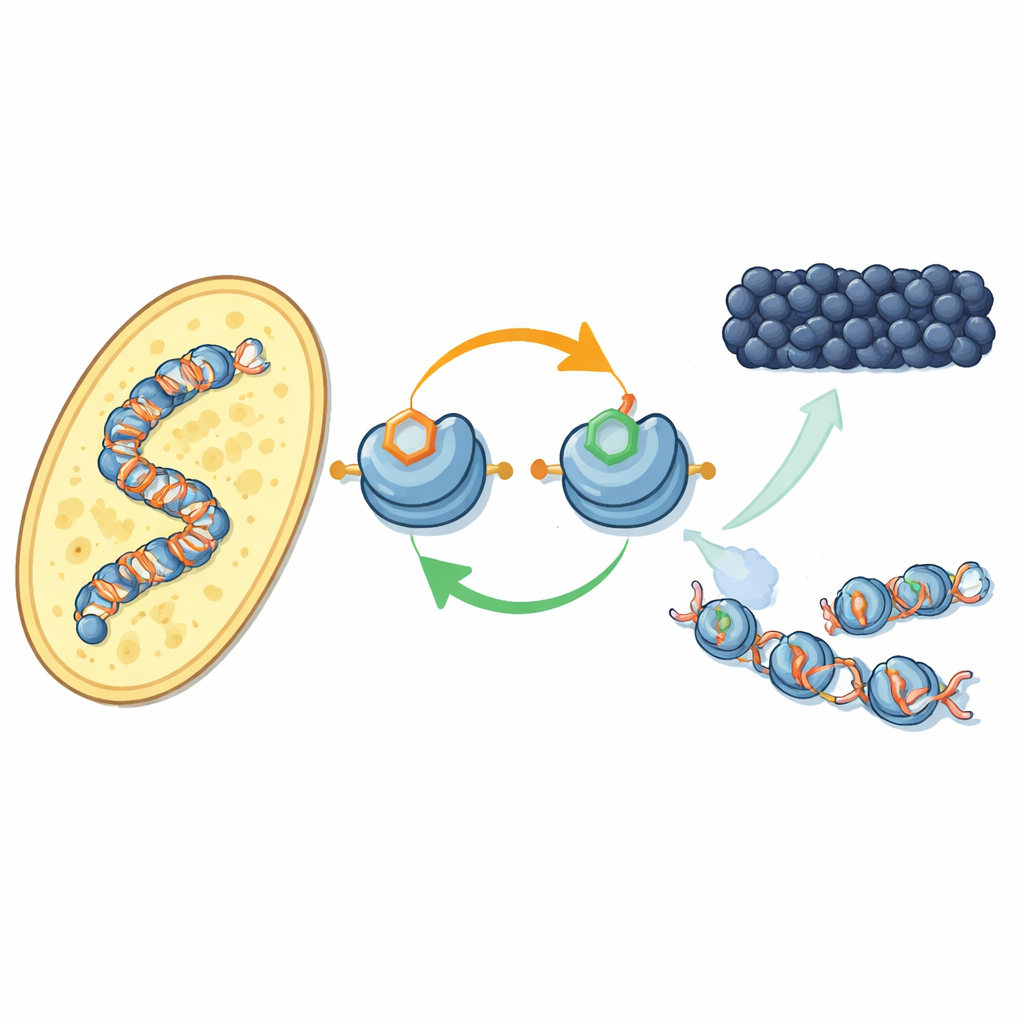
En ny återkopplingsslinga på DNA-packningsproteiner
Författarna fokuserar på hur specifika kemiska märkningar på histonproteiner — spolarna som DNA är lindat runt — samarbetar. I fissionsjäst lägger ett enzym kallat Clr4 till en metylgrupp på en position på histon H3 (känd som H3K9), ett kännetecken för heterokromatin. Samma enzym ingår också i ett större komplex, CLRC, som fäster en liten ubiquitin-molekyl på en annan position på H3 (H3K14). Med renade komponenter visar forskarna att tillsats av ubiquitin på H3K14 dramatiskt ökar Clr4:s förmåga att metylera H3K9 på fullständiga nukleosomer och därigenom övervinner en annars hämmande effekt av DNA. Denna stimulerande effekt är mycket specifik: andra ubiquitin-märkningar på olika histonpositioner ger inte samma påverkan.
Två markeringar som upprätthåller varandra
På levande jästceller använde teamet en nyligen utvecklad antikropp för att kartlägga var ubiquitin-taggen på H3K14 förekommer i genomet. Den sitter nästan exakt där H3K9-metylering gör det — vid centromerer, telomerer och en tyst region för parnings-typ — klassiska heterokromatinterritorier. När de inaktiverade antingen metylmarkeringen eller enzymerna som sätter den försvann också ubiquitin-markeringen, och vice versa. Båda märkningarna behövs för att hålla CLRC-komplexet förankrat på kromatin. Tillsammans visar detta en positiv återkopplingsslinga: ubiquitin-markeringen ökar metyleringen, och paret av markeringar stabiliserar just det komplex som sätter dem, vilket hjälper det tysta tillståndet att sprida sig till närliggande nukleosomer och återställas efter DNA-replikation.
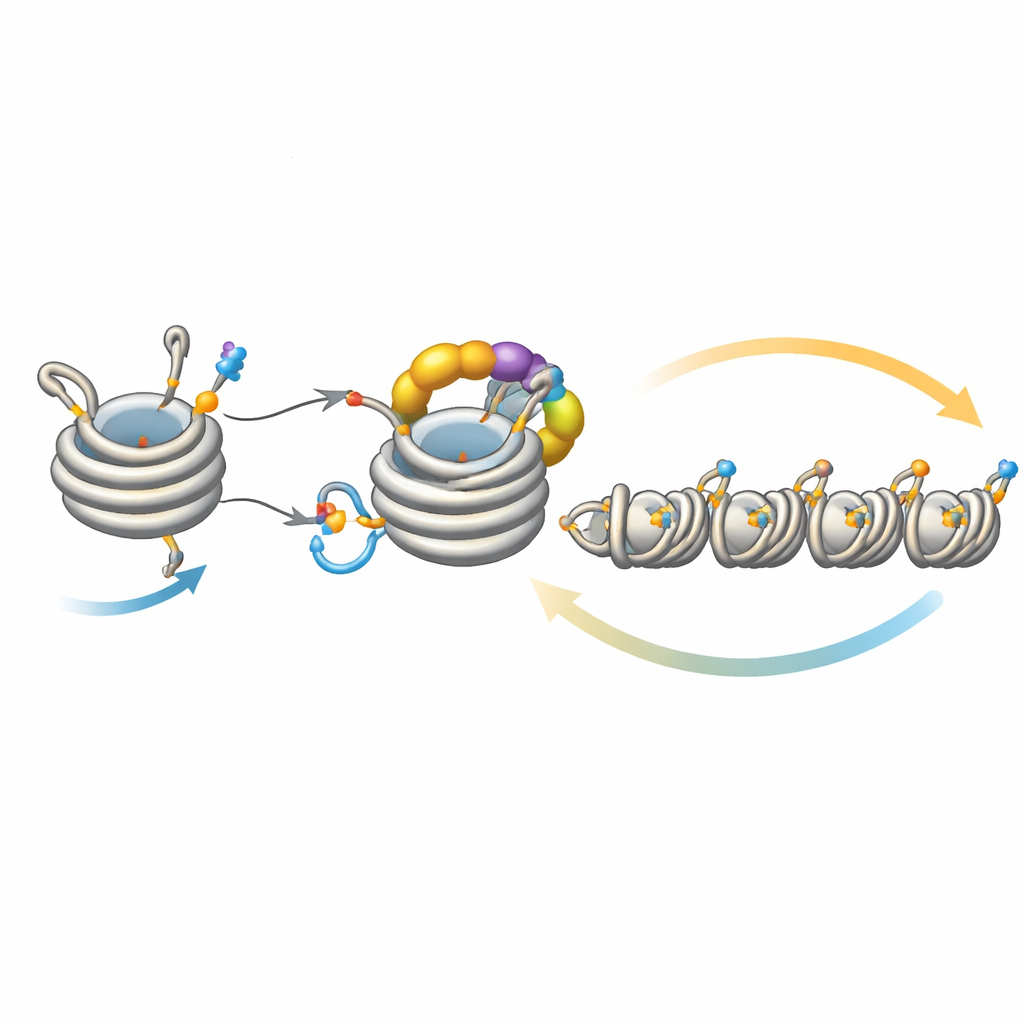
Finjustering av tystnadens spridning
Denne slinga arbetar inte isolerat. Ett deacetylasenzym som heter Clr3 förbereder histon H3 för ubiquitinering genom att ta bort en acetylgrupp på samma plats, H3K14; när detta steg är nedsatt försvagas återkopplingen och heterokromatin har svårt att sprida sig eller ärvas, särskilt i regioner långt från de ursprungliga nukleationsställena. Omvänt agerar flera enzymer som bromsar. Mst2 lägger till acetylgruppen som blockerar ubiquitineringen, och Epe1 tar bort metylmarkeringen på H3K9. Att avlägsna dessa bromsar gör att heterokromatin sprider sig för långt, släcker viktiga gener och stressar cellerna. Under sådan stress utlöser cellerna ett adaptivt svar som minskar mängden Clr4 och dämpar den överaktiva återkopplingsslingan. Genetiska experiment visar att förstärkning eller försvagning av olika delar av denna krets kan rädda eller eliminera defekter i spridning och arv, vilket understryker hur ömt systemet är balanserat.
Konsekvenser bortom jästceller
Tillsammans visar resultaten att heterokromatin inte upprätthålls av en enda svag "läs-skriv"-reaktion, utan av ett integrerat kretslopp av ubiquitinering, deacetylation och metylering på histonproteiner. I fissionsjäst skapar denna H3K14ub–H3K9me3-återkopplingsslinga en tvåstabil strömbrytare: när en region korsar en tröskel av dessa markeringar tenderar den att förbli tyst och sprida sig, medan regioner som inte når tröskeln aldrig fullt ut blir heterokromatin. Eftersom både dessa histonmarkeringar och många av de ingående enzymerna är bevarade hos däggdjur kan en liknande logik bidra till att upprätthålla stabila men flexibla mönster av genrepression i humana celler. Förståelsen av denna krets kan så småningom informera nya sätt att modulera kromatin — till exempel för att stabilisera genomet i åldrande celler eller för att selektivt återaktivera tysta regioner vid cancerterapi.
Citering: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
Nyckelord: heterokromatin, histonmodifiering, epigenetiskt arv, kromatinåterkopplingsslinga, fissionsjäst