Clear Sky Science · fr
Un circuit de rétroaction H3K14ub-H3K9me3 régit la propagation et l’héritage de l’hétérochromatine chez la levure de fission
Pourquoi cet interrupteur microscopique compte
À l’intérieur de chacune de nos cellules, de longues molécules d’ADN sont soigneusement compactées et gérées pour que les bons gènes s’activent ou se désactivent au bon moment. Une part importante de ce système de contrôle est l’« hétérochromatine », l’ADN fortement compacté qui maintient certaines portions du génome silencieuses et stables. Cette étude, réalisée chez la levure de fission, met au jour un circuit moléculaire de rétroaction caché qui aide l’hétérochromatine à se propager le long des chromosomes et à persister lors des divisions cellulaires. Parce que des systèmes semblables existent chez l’humain, comprendre ce circuit fournit des indices sur la façon dont les cellules préservent la stabilité du génome et comment la défaillance de ce mécanisme peut contribuer aux maladies, voire au cancer.
Un quartier silencieux du génome
Toute l’ADN n’est pas également actif. Certaines régions forment de l’hétérochromatine, où les gènes sont pour la plupart éteints. Ces étendues silencieuses protègent le génome en empêchant les séquences répétées de mal fonctionner, en limitant les recombinaisons nuisibles et en façonnant les programmes d’expression génique. L’hétérochromatine comporte trois étapes clés : elle démarre à des sites spéciaux de « nucléation », se propage dans les régions voisines, puis est transmise au cours des divisions cellulaires. Si les signaux d’amorçage sont relativement bien compris, les scientifiques se sont longtemps interrogés sur la manière dont l’état silencieux se propage et perdure de façon fiable, d’autant que les modèles précédents reposaient sur une marque d’auto-renforcement relativement faible sur une histone.
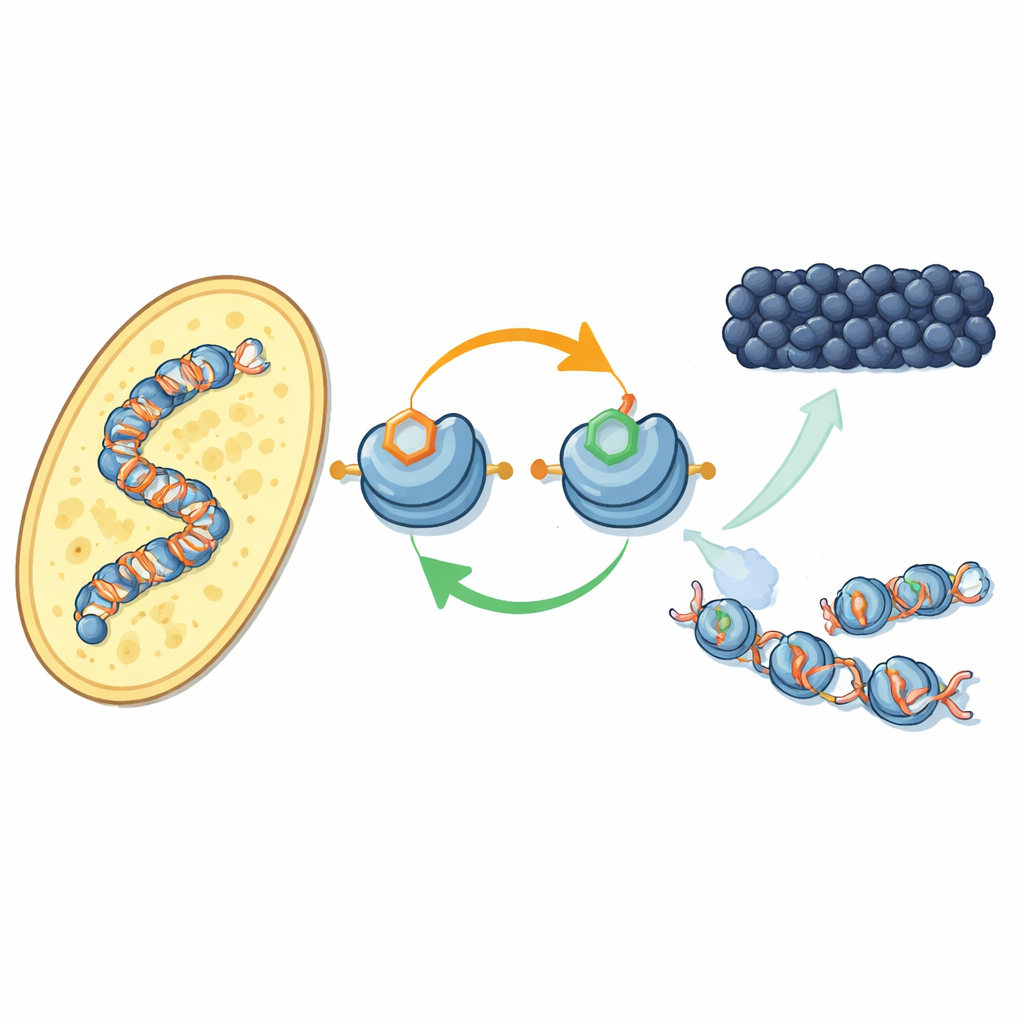
Une nouvelle boucle de rétroaction sur les protéines d’emballage de l’ADN
Les auteurs se concentrent sur la manière dont des marques chimiques spécifiques sur les protéines histones — les « bobines » autour desquelles l’ADN est enroulé — coopèrent. Chez la levure de fission, une enzyme appelée Clr4 ajoute un groupement méthyle à une position de l’histone H3 (connue sous le nom H3K9), marque caractéristique de l’hétérochromatine. La même enzyme fait aussi partie d’un complexe plus vaste, CLRC, qui ajoute un petit marqueur ubiquitine à une autre position de H3 (H3K14). Avec des composants purifiés, les chercheurs montrent que l’addition d’ubiquitine sur H3K14 augmente de manière spectaculaire la capacité de Clr4 à méthyler H3K9 sur des nucléosomes complets, surmontant un effet inhibiteur exercé par l’ADN. Cet effet stimulant est très spécifique : d’autres ubiquitinations à d’autres positions d’histones n’ont pas le même impact.
Deux marques qui se soutiennent mutuellement
En se tournant vers des cellules de levure vivantes, l’équipe a utilisé un anticorps récemment développé pour cartographier l’emplacement de la marque ubiquitine H3K14 à travers le génome. Elle se situe presque exactement là où apparaît la méthylation de H3K9 — aux centromères, aux télomères et dans une région silencieuse de type accouplement — des territoires classiques d’hétérochromatine. Lorsque les auteurs inactivaient soit la marque méthyle, soit les enzymes qui la déposent, la marque ubiquitine disparaissait aussi, et inversement. Les deux marques sont nécessaires pour ancrer le complexe CLRC sur la chromatine. Ensemble, cela révèle une boucle de rétroaction positive : la marque ubiquitine stimule la méthylation, et la paire de marques stabilise le complexe qui les place, permettant à l’état silencieux de se propager aux nucléosomes voisins et d’être restauré après la réplication de l’ADN.
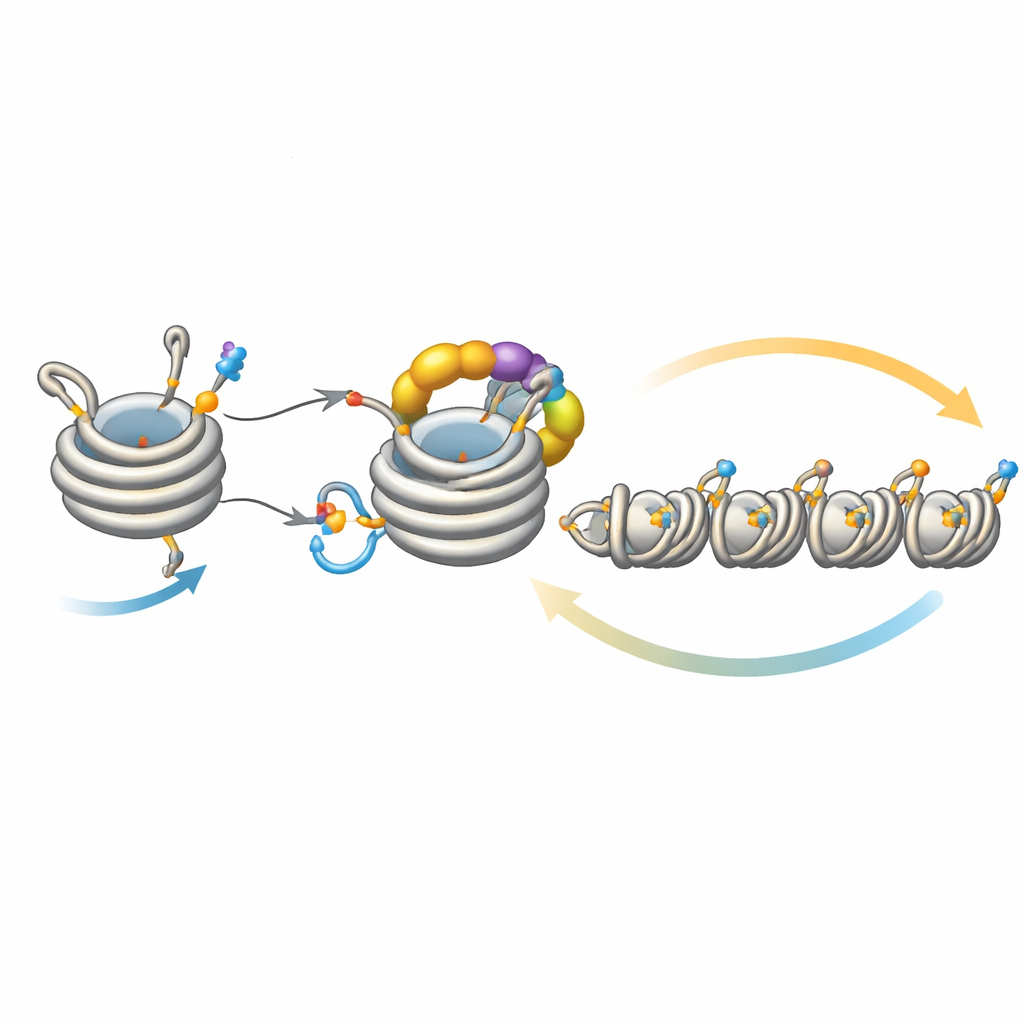
Ajuster finement la propagation du silence
Cette boucle ne fonctionne pas isolément. Une déacétylase appelée Clr3 prépare l’histone H3 à l’ubiquitination en retirant un groupe acétyle à ce même site, H3K14 ; lorsque cette étape est perturbée, la rétroaction s’affaiblit et l’hétérochromatine peine à se propager ou à être héritée, en particulier dans des régions éloignées des sites de nucléation initiaux. À l’inverse, plusieurs enzymes font office de freins. Mst2 ajoute le groupe acétyle qui empêche l’ubiquitination, et Epe1 élimine la marque méthyle sur H3K9. Supprimer ces freins permet à l’hétérochromatine de trop se propager, en silencant des gènes essentiels et en stressant les cellules. Sous un tel stress, les cellules déclenchent une réponse adaptative qui réduit la quantité de Clr4, diminuant la boucle de rétroaction hyperactive. Des expériences génétiques montrent que renforcer ou affaiblir différentes branches de ce circuit peut sauver ou abolir des défauts de propagation et d’héritage, soulignant l’équilibre délicat du système.
Des implications au-delà des cellules de levure
Dans l’ensemble, les résultats révèlent que l’hétérochromatine n’est pas maintenue par une seule réaction de « lecture-écriture » faible, mais par un circuit intégré d’ubiquitination, de déacétylation et de méthylation sur les histones. Chez la levure de fission, cette boucle de rétroaction H3K14ub–H3K9me3 crée un commutateur bistable : une fois qu’une région dépasse un seuil de ces marques, elle a tendance à rester silencieuse et à se propager, tandis que les régions qui ne l’atteignent pas ne s’engagent jamais complètement dans l’hétérochromatine. Parce que ces deux marques d’histones et bon nombre des enzymes impliquées sont conservées chez les mammifères, une logique similaire pourrait aider à maintenir des motifs de répression génique stables mais flexibles dans les cellules humaines. Comprendre ce circuit pourrait éventuellement permettre de nouvelles manières de moduler la chromatine — par exemple, pour stabiliser le génome dans les cellules vieillissantes ou pour réveiller sélectivement des régions silencieuses lors de thérapies anticancéreuses.
Citation: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
Mots-clés: hétérochromatine, modification des histones, héritage épigénétique, boucle de rétroaction chromatinienne, levure de fission