Clear Sky Science · pt
Um circuito de realimentação H3K14ub-H3K9me3 governa a propagação e a herança da heterocromatina em levedura de fissão
Por que esse interruptor microscópico importa
Dentro de cada uma de nossas células, longas fitas de DNA são cuidadosamente empacotadas e gerenciadas para que os genes certos sejam ativados ou desativados no momento adequado. Uma parte importante desse sistema de controle é a "heterocromatina", DNA densamente compactado que mantém determinados trechos do genoma silenciosos e estáveis. Este estudo, realizado em levedura de fissão, desvenda um circuito molecular de realimentação oculto que ajuda a heterocromatina a se espalhar ao longo dos cromossomos e a persistir quando as células se dividem. Como sistemas semelhantes existem em humanos, entender esse circuito oferece pistas sobre como as células mantêm a estabilidade do genoma e como sua falha pode contribuir para doenças e até câncer.
Um bairro silencioso no genoma
Nem todo DNA é igualmente ativo. Algumas regiões formam heterocromatina, onde os genes ficam em grande parte desligados. Esses trechos silenciosos protegem o genoma ao evitar que DNA repetitivo cause problemas, limitar recombinações prejudiciais e moldar programas de expressão gênica. A heterocromatina tem três fases-chave: começa em locais especiais de "nucleação", espalha-se para as regiões vizinhas e então é transmitida através das divisões celulares. Embora os sinais de início sejam relativamente bem compreendidos, os cientistas há muito se perguntam como o estado silencioso se espalha e perdura de forma confiável, especialmente considerando que modelos anteriores se apoiavam em uma marca auto-reforçadora relativamente fraca em uma única histona.
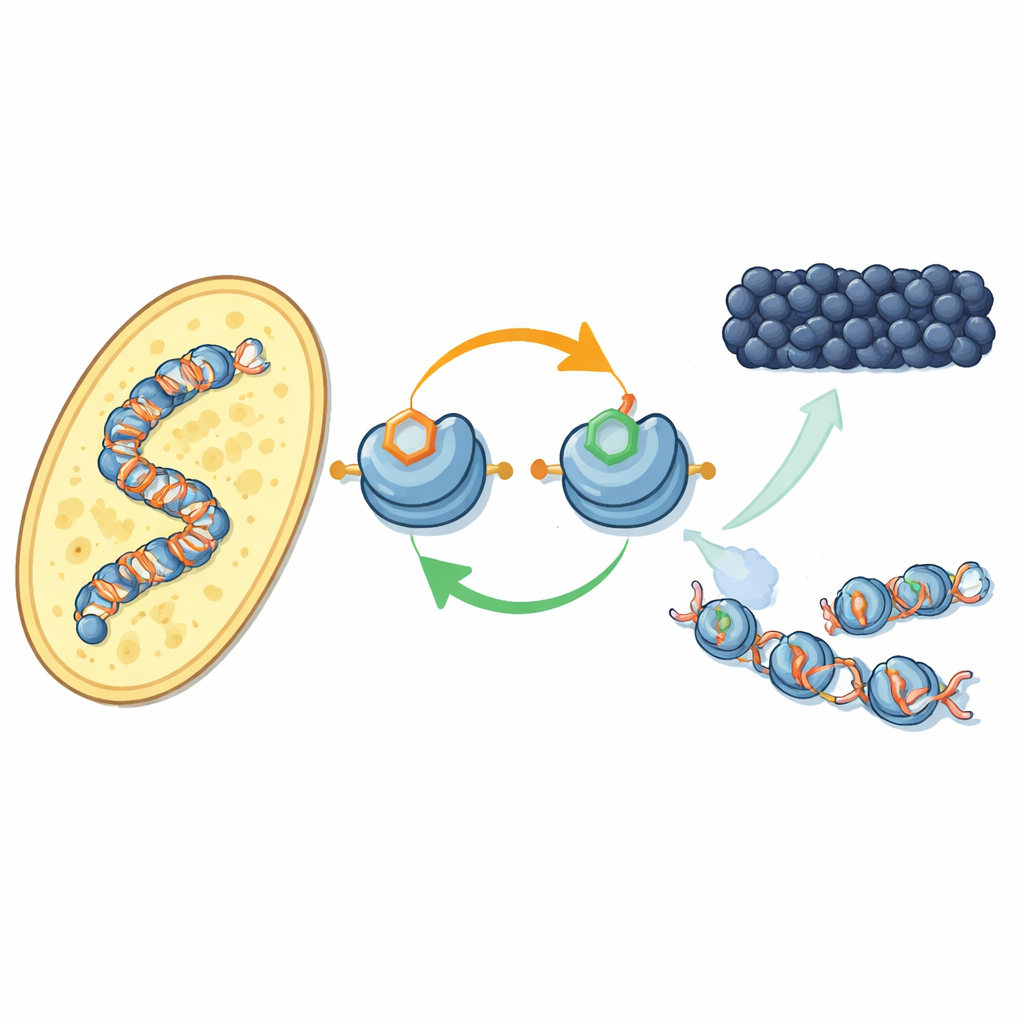
Um novo circuito de realimentação nas proteínas de empacotamento do DNA
Os autores concentram-se em como marcas químicas específicas nas histonas — os carretéis em torno dos quais o DNA é enrolado — atuam em conjunto. Na levedura de fissão, uma enzima chamada Clr4 adiciona uma marca de metil no ponto H3K9 da histona H3, um marco da heterocromatina. A mesma enzima também integra um complexo maior, o CLRC, que acrescenta uma pequena marca de ubiquitina em outro ponto da H3 (H3K14). Usando componentes purificados, os pesquisadores mostram que a adição de ubiquitina em H3K14 aumenta dramaticamente a capacidade de Clr4 de metilar H3K9 em nucleossomos completos, superando um efeito inibitório imposto pelo DNA. Esse efeito estimulatório é altamente específico: outras marcas de ubiquitina em posições diferentes das histonas não produzem o mesmo impacto.
Duas marcas que se sustentam mutuamente
Em células de levedura vivas, a equipe utilizou um anticorpo recém-desenvolvido para mapear onde a ubiquitinação em H3K14 aparece no genoma. Ela localiza-se quase exatamente onde a metilação de H3K9 ocorre — nos centrômeros, telômeros e numa região silenciosa de tipo de acasalamento — territórios clássicos de heterocromatina. Quando desativaram ou a marca de metilação ou as enzimas que a colocam, a marca de ubiquitina também desapareceu, e vice-versa. Ambas as marcas são necessárias para manter o complexo CLRC ancorado na cromatina. Conjuntamente, isso revela um loop de realimentação positivo: a ubiquitina aumenta a metilação, e o par de marcas estabiliza o próprio complexo que as deposita, ajudando o estado silencioso a se espalhar por nucleossomos vizinhos e a ser restaurado após a replicação do DNA.
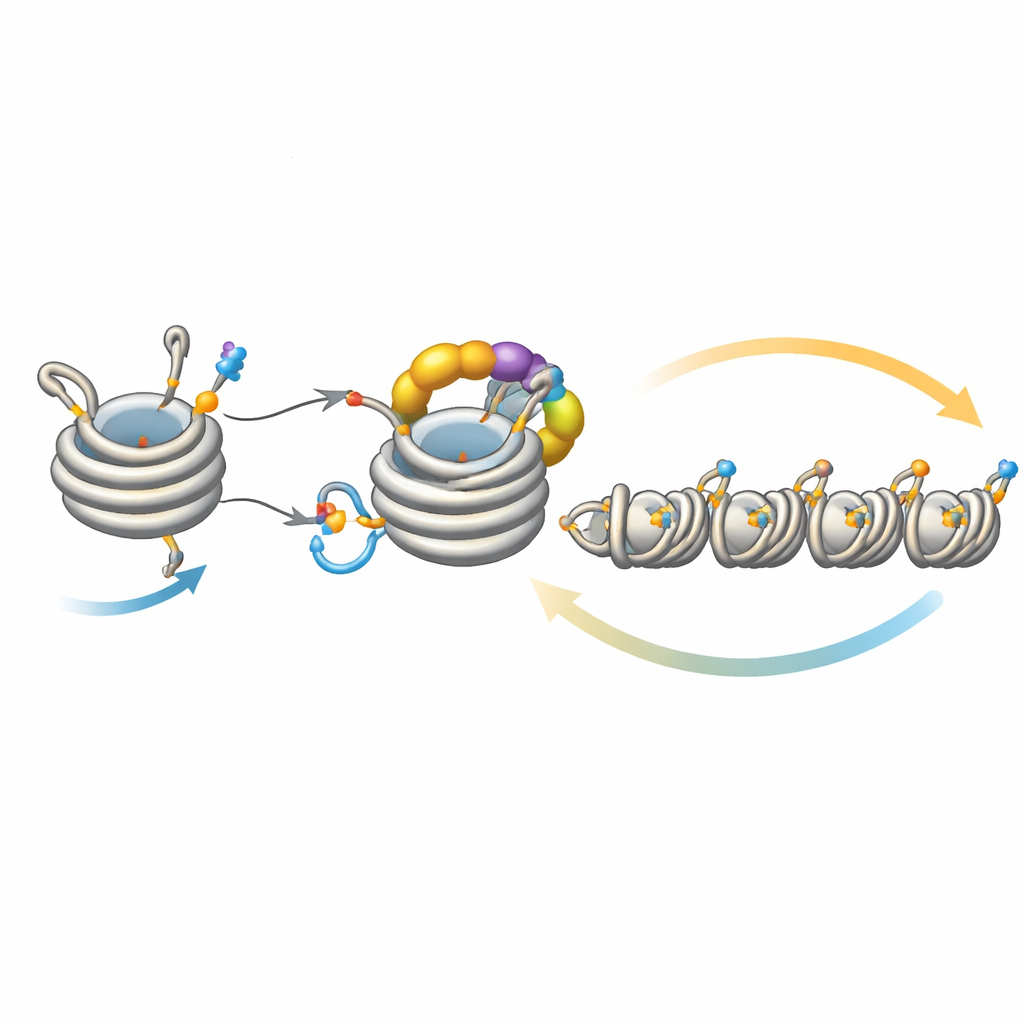
Ajustando finamente a propagação do silêncio
Esse circuito não funciona isoladamente. Uma desacetilase chamada Clr3 prepara a histona H3 para a ubiquitinação ao remover um grupo acetil no mesmo sítio, H3K14; quando essa etapa é prejudicada, a realimentação enfraquece e a heterocromatina tem dificuldade de se espalhar ou ser herdada, especialmente em regiões distantes dos sítios de nucleação originais. Em contrapartida, várias enzimas atuam como freios. Mst2 adiciona o grupo acetil que bloqueia a ubiquitinação, e Epe1 remove a marca de metil em H3K9. Eliminar esses freios faz a heterocromatina se espalhar demais, silenciando genes essenciais e estressando as células. Sob esse estresse, as células acionam uma resposta adaptativa que reduz a quantidade de Clr4, diminuindo o loop de realimentação hiperativo. Experimentos genéticos mostram que fortalecer ou enfraquecer diferentes braços desse circuito pode resgatar ou abolir defeitos na propagação e herança, ressaltando o quão delicadamente equilibrado o sistema é.
Implicações além das células de levedura
No conjunto, os achados revelam que a heterocromatina não é mantida por uma única reação "ler-escrever" fraca, mas por um circuito integrado de ubiquitinação, desacetilação e metilação nas histonas. Na levedura de fissão, esse loop de realimentação H3K14ub–H3K9me3 cria um interruptor biestável: uma vez que uma região ultrapassa um limiar dessas marcas, tende a permanecer silenciosa e a se espalhar, enquanto regiões que não atingem esse limiar nunca se comprometem totalmente com a heterocromatina. Como ambas as marcas de histona e muitas das enzimas envolvidas são conservadas em mamíferos, uma lógica similar pode ajudar a manter padrões estáveis, porém flexíveis, de repressão gênica em células humanas. Compreender essa circuitaria pode, no futuro, informar novas maneiras de modular a cromatina — por exemplo, para estabilizar o genoma em células envelhecidas ou para reativar seletivamente regiões silenciosas durante terapias contra o câncer.
Citação: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
Palavras-chave: heterocromatina, modificação de histonas, herança epigenética, loop de feedback da cromatina, levedura de fissão