Clear Sky Science · de
Ein H3K14ub–H3K9me3-Feedbackkreis steuert die Ausbreitung und Vererbung von Heterochromatin in Spalthefe
Warum dieser mikroskopische Schalter wichtig ist
In jeder unserer Zellen sind lange DNA-Stränge sorgfältig verpackt und reguliert, damit die richtigen Gene zur richtigen Zeit ein- oder ausgeschaltet werden. Ein wesentlicher Teil dieses Kontrollsystems ist das "Heterochromatin" — dicht gepackte DNA, die bestimmte Abschnitte des Genoms stumm und stabil hält. Diese Studie, durchgeführt in der Spalthefe, enthüllt einen verborgenen molekularen Feedbackkreis, der die Ausbreitung von Heterochromatin entlang der Chromosomen und dessen Persistenz bei Zellteilungen fördert. Da ähnliche Systeme auch im Menschen existieren, liefert das Verständnis dieses Kreises Hinweise darauf, wie Zellen die Genomstabilität aufrechterhalten und wie das Versagen dieser Mechanismen zu Krankheiten und sogar Krebs beitragen kann.
Eine ruhige Nachbarschaft im Genom
Nicht alle DNA ist gleichermaßen aktiv. Manche Regionen bilden Heterochromatin, in dem Gene weitgehend abgeschaltet sind. Diese stillen Abschnitte schützen das Genom, indem sie repetitiven DNA-Sequenzen Einhalt gebieten, schädliche Rekombinationen begrenzen und Expressionsprogramme von Genen mitgestalten. Heterochromatin durchläuft drei wesentliche Phasen: Es beginnt an speziellen "Nukleations"-Stellen, breitet sich in benachbarte Regionen aus und wird dann durch Zellteilungen weitergegeben. Während die anfänglichen Signale relativ gut verstanden sind, rätseln Wissenschaftler seit langem darüber, wie der stille Zustand zuverlässig ausgedehnt und erhalten bleibt — zumal frühere Modelle sich auf eine vergleichsweise schwache sich selbst verstärkende Markierung an einem Histon stützen.
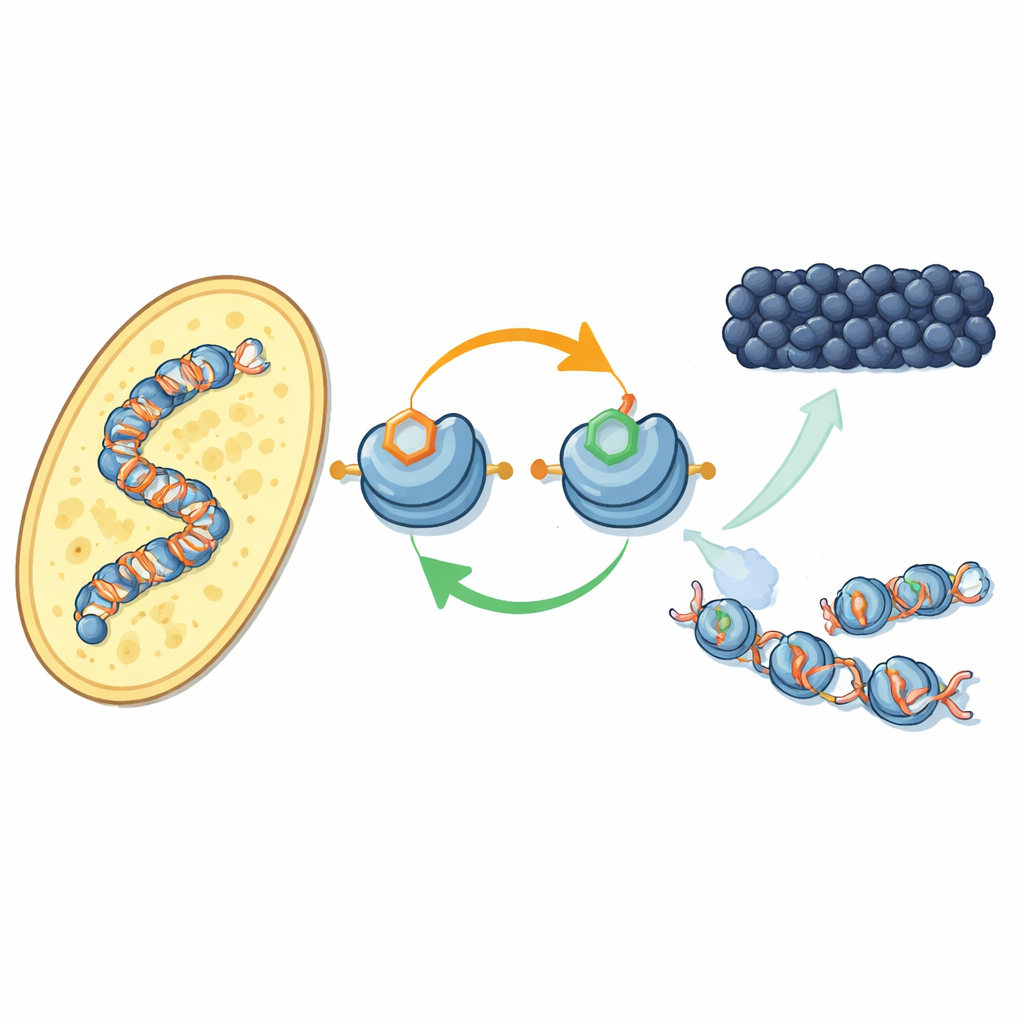
Eine neue Rückkopplung an den DNA-Packproteinen
Die Autoren konzentrieren sich darauf, wie bestimmte chemische Markierungen an Histonproteinen — den Spulen, um die die DNA gewickelt ist — zusammenwirken. In der Spalthefe fügt ein Enzym namens Clr4 eine Methylmarke an einer Position des Histons H3 hinzu (bekannt als H3K9), ein Kennzeichen von Heterochromatin. Dasselbe Enzym ist außerdem Bestandteil eines größeren Komplexes, CLRC, der an einer anderen Position von H3 (H3K14) ein kleines Ubiquitin-Tag anbringt. Mit gereinigten Komponenten zeigen die Forscher, dass die Ubiquitinierung von H3K14 die Fähigkeit von Clr4, H3K9 auf vollständigen Nukleosomen zu methylieren, deutlich steigert und damit eine sonst hemmende Wirkung der DNA überwindet. Dieser stimulierende Effekt ist hochspezifisch: Andere Ubiquitinanhängungen an anderen Histonpositionen haben nicht dieselbe Wirkung.
Zwei Markierungen, die einander stützen
In lebenden Hefezellen verwendete das Team einen neu entwickelten Antikörper, um zu kartieren, wo das H3K14-Ubiquitin-Tag im Genom zu finden ist. Es liegt nahezu exakt dort, wo auch H3K9-Methylierung auftritt — an Centromeren, Telomeren und in einer stillen Region des Paarungstyps — klassische Heterochromatin-Territorien. Wenn sie entweder die Methylmarke oder die Enzyme, die sie anbringen, ausschalteten, verschwand auch das Ubiquitin-Tag, und umgekehrt. Beide Markierungen sind notwendig, damit der CLRC-Komplex auf dem Chromatin verankert bleibt. Zusammen ergibt sich ein positiver Feedbackkreis: Das Ubiquitin-Tag fördert die Methylierung, und das Paar von Markierungen stabilisiert den Komplex, der sie setzt, wodurch der stille Zustand auf benachbarte Nukleosomen übertragen und nach der DNA-Replikation wiederhergestellt wird.
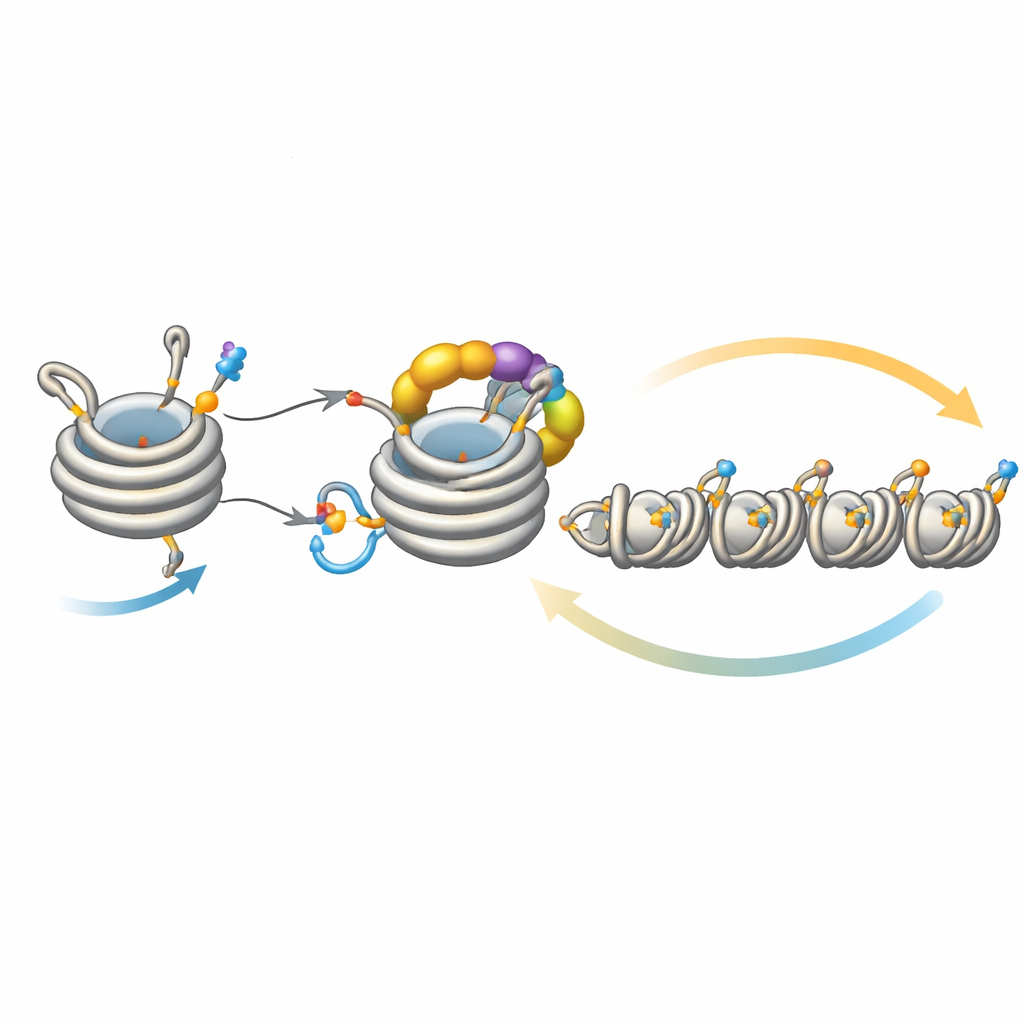
Die Ausbreitung der Stille feinabstimmen
Dieser Kreis wirkt nicht isoliert. Ein Deacetylase-Enzym namens Clr3 bereitet Histon H3 für die Ubiquitinierung vor, indem es eine Acetylgruppe an derselben Stelle, H3K14, entfernt; ist dieser Schritt gestört, schwächt das Feedback und Heterochromatin hat Schwierigkeiten, sich auszubreiten oder vererbt zu werden, insbesondere in Regionen weit entfernt von den ursprünglichen Nukleationsstellen. Umgekehrt wirken mehrere Enzyme als Bremsen. Mst2 fügt die Acetylgruppe hinzu, die die Ubiquitinierung blockiert, und Epe1 entfernt die Methylmarke an H3K9. Das Entfernen dieser Bremsen lässt Heterochromatin zu weit ausbreiten, schaltet essentielle Gene stumm und belastet die Zellen. Unter diesem Stress lösen die Zellen eine Anpassungsreaktion aus, die die Menge an Clr4 reduziert und damit den überaktiven Feedbackkreis dämpft. Genetische Experimente zeigen, dass das Verstärken oder Abschwächen verschiedener Schaltstellen dieses Kreises Defekte in Ausbreitung und Vererbung beheben oder hervorrufen kann — ein Hinweis auf die feine Balance dieses Systems.
Folgen über Hefe hinaus
Die Ergebnisse zeigen zusammen, dass Heterochromatin nicht durch eine einzige schwache "Lese-Schreib"-Reaktion erhalten bleibt, sondern durch einen integrierten Schaltkreis aus Ubiquitinierung, Deacetylierung und Methylierung an Histonproteinen. In der Spalthefe erzeugt dieser H3K14ub–H3K9me3-Feedbackkreis einen bistabilen Schalter: Sobald eine Region eine Schwelle dieser Markierungen überschreitet, bleibt sie tendenziell stumm und breitet sich aus, während Regionen, die diese Schwelle nicht erreichen, nie vollständig in den Heterochromatin-Zustand übergehen. Da beide Histonmarken und viele der beteiligten Enzyme auch in Säugetieren konserviert sind, könnte eine ähnliche Logik dazu beitragen, stabile und zugleich flexible Muster der Genrepression in menschlichen Zellen zu erhalten. Das Verständnis dieser Schaltkreise könnte langfristig neue Wege eröffnen, Chromatin zu modulieren — etwa um das Genom in alternsbedingten Zellen zu stabilisieren oder stille Regionen gezielt während einer Krebstherapie wieder zu aktivieren.
Zitation: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
Schlüsselwörter: Heterochromatin, Histonmodifikation, epigenetische Vererbung, Chromatin-Feedbackschleife, Spalthefe