Clear Sky Science · he
מעגל משוב H3K14ub–H3K9me3 שולט בהתפשטות ובהורשה של הטרוכרומטין בשמרי פיצול
מדוע המתג המיקרוסקופי הזה חשוב
בתוך כל אחת מהתאים שלנו, גדילי DNA ארוכים נארזים ומנוהלים בקפידה כדי שהגנים המתאימים יידלקו או ייכנסו לדי-אקטיבציה בזמנים הנכונים. חלק מרכזי במערכת הבקרה הזו הוא "הטרוכרומטין" — DNA דחוס מאוד השומר על מקטעים מסוימים של הגנום בשקט וביציבות. המחקר הזה, שנערך בשמרי פיצול, חושף מעגל משוב מולקולרי נסתר שעוזר להטרוכרומטין להתפשט לאורך הכרומוזומים ולהישאר קיים כשתאים מתחלקים. מכיוון שמערכות דומות קיימות בבני אדם, הבנת המעגל הזה מעניקה רמזים כיצד תאים שומרים על יציבות הגנום וכיצד כשלים בו עלולים לתרום למחלה ואפילו לסרטן.
שכונה שקטה בגנום
לא כל ה-DNA פעיל באותה מידה. אזורים מסוימים יוצרים הטרוכרומטין, שבו גנים ברובם מושתקים. מאווררים אלו מגנים על הגנום על ידי מניעת התנהגות לקויה של רצפים חוזרים, הגבלת רקומבינציה מזיקה ועיצוב תכניות ביטוי גנים. להטרוכרומטין שלוש שלבים מרכזיים: הוא מתחיל באתרי "הנבטה" מיוחדים, מתפשט לאזורים שכנים, ואז מועבר דרך חלוקות תאים. בעוד שהאותות ההתחלתיים מובנים יחסית טוב, מדענים התלבטו זמן רב כיצד המצב השקט מתפשט ומתקיים בצורה אמינה, במיוחד בהתחשב בעובדה שמודלים קודמים נשענו על סימן עצמי-מחזק חלש יחסית בחלבון היסטון אחד.
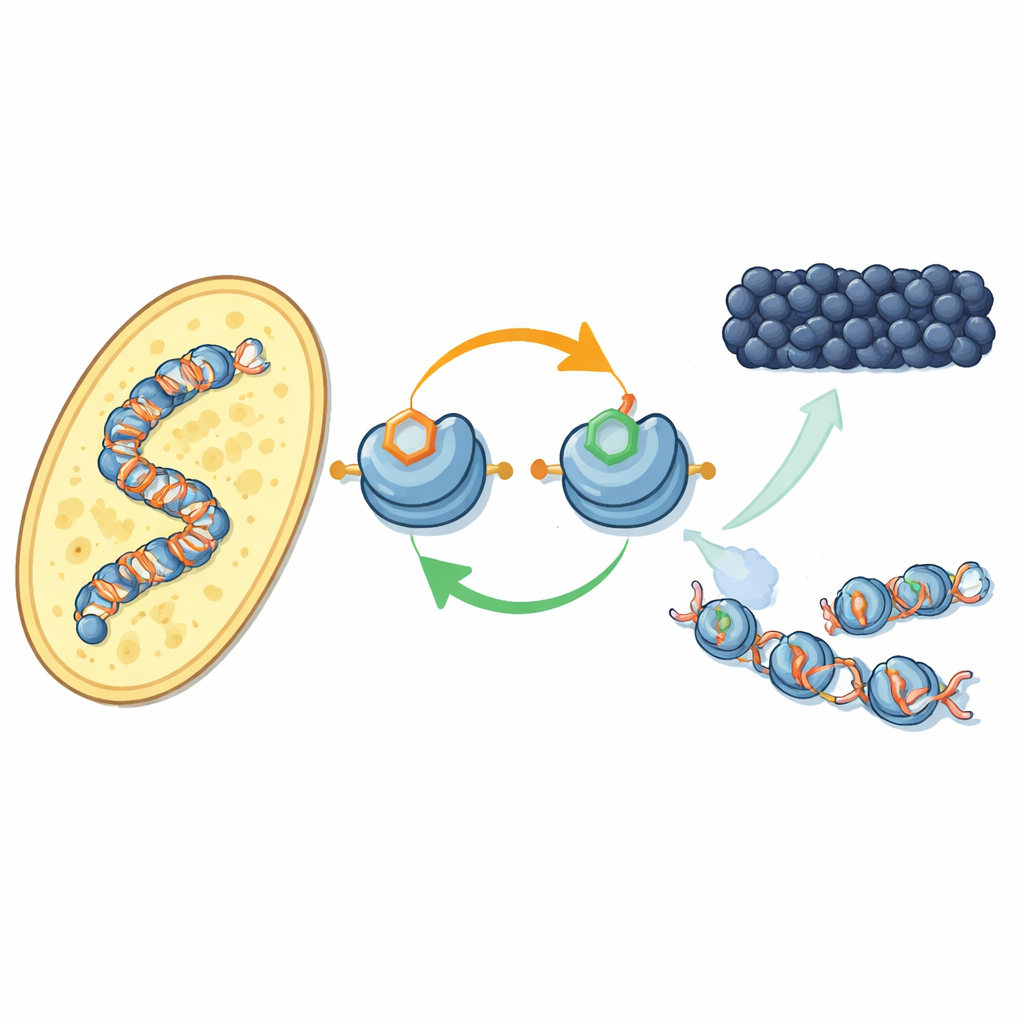
לולאת משוב חדשה על חלבוני אריזת ה-DNA
המחברים מתמקדים באופן שבו תגיות כימיות ספציפיות על חלבוני ההיסטון — הסלילים שסביבם ה-DNA נכרך — פועלות יחד. בשמרי פיצול, אנזים בשם Clr4 מוסיף תג מתיל בעמדה מסוימת על היסטון H3 (ידוע כ-H3K9), סימן היכר של הטרוכרומטין. אותו אנזים גם נמצא בקומפלקס גדול יותר, CLRC, שמצמיד תג יוביקוויטין קטן בעמדה אחרת על H3 (H3K14). בעבודה עם רכיבים מטוהרים, החוקרים מראים שהוספת יוביקוויטין ב-H3K14 מגבירה דרמטית את יכולתו של Clr4 למתל את H3K9 על נוקלאוזומים שלמים, ומתגברת על אפקט מעכב של ה-DNA. ההשפעה המעוררת הזו היא מאוד ספציפית: תגיות יוביקוויטין אחרות בעמדות שונות בהיסטון אינן משפיעות באותה מידה.
שני סימנים שמחזיקים זה את זה
בעבודה על תאים חיים של שמרים, הצוות השתמש בנוגדן חדש למיפוי היכן תג יוביקוויטין H3K14 מופיע בגנום. הוא נמצא כמעט בדיוק במקומות שבהם מתילציית H3K9 מתרחשת — במרכזי כרומוזומים, בטלומרים ובאזור שקט של סוג הזיווג — שטחי הטרוכרומטין הקלאסיים. כשכיבו או השביתו את סימן המתיל או את האנזימים שמציבים אותו, תג היוביקוויטין נעלם גם כן, ולהפך. שני הסימנים נחוצים כדי לשמר את קומפלקס CLRC קשור לכרומטין. ביחד, ממצאים אלה חושפים לולאת משוב חיובית: תג היוביקוויטין מעודד מתילציה, ודו-סימן זה מייצב את הקומפלקס שמציב אותם, מה שעוזר למצב השקט להתפשט לנוקלאוזומים שכנים ולהישתקם אחרי שכפול ה-DNA.
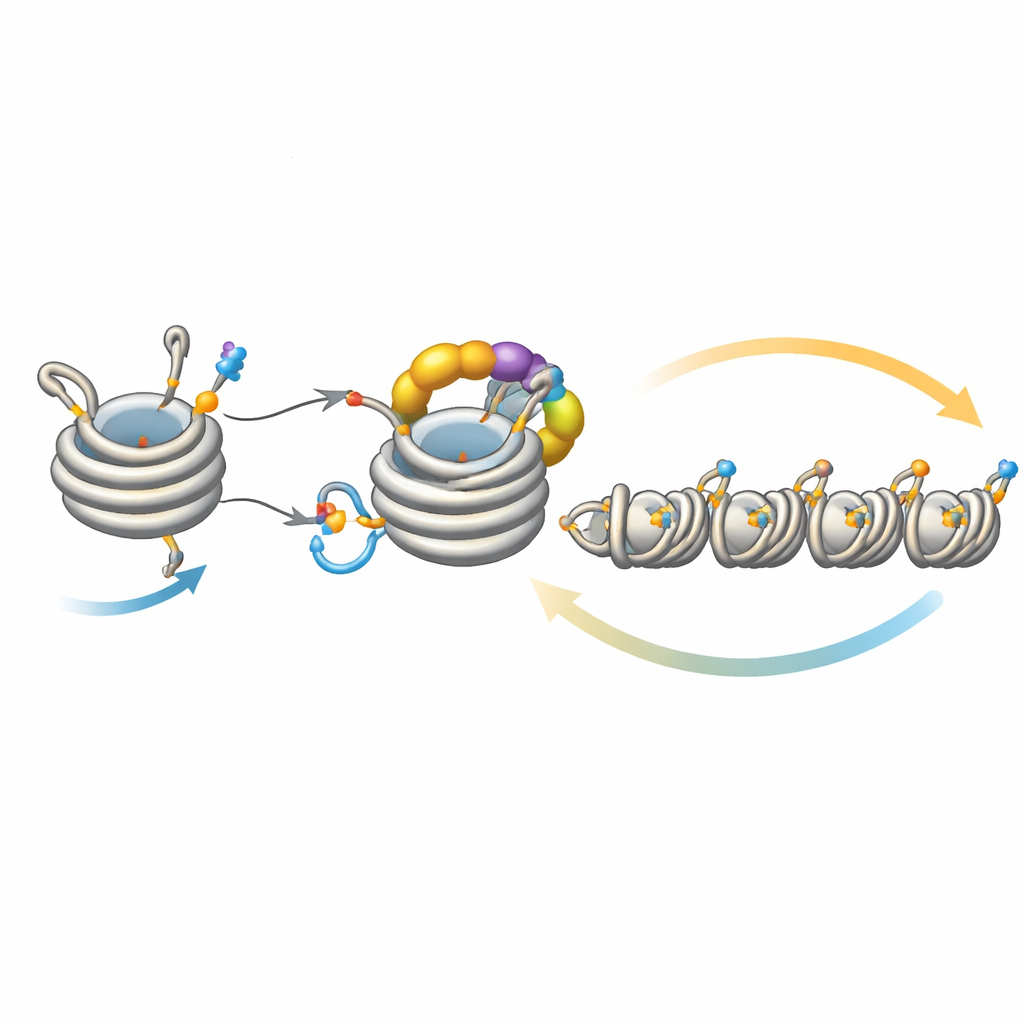
כוונון עדין של התפשטות השקט
הלולאה הזו לא פועלת בבידוד. אנזים דה-אצטילאז בשם Clr3 מכין את H3 לאוביקויטינציה על ידי הסרת קבוצת אצטיל באותו אתר, H3K14; כאשר שלב זה מופרע, המשוב נחלש והטרוכרומטין מתקשה להתפשט או להיות מוטווה לדורות הבאים, במיוחד באזורים רחוקים מאתרי ההנבטה המקוריים. מצד שני, כמה אנזימים פועלים כבלמים. Mst2 מוסיף את קבוצת האצטיל שחוסמת את האוביקויטינציה, ו-Epe1 מסיר את סימן המתיל על H3K9. הסרת הבלמים האלו גורמת להתרחבות מופרזת של הטרוכרומטין, מה שמושתק גנים חיוניים ומציב לחץ על התאים. במצב של לחץ כזה התאים מפעילים תגובה אדפטיבית שמפחיתה את כמות Clr4, ובכך מדכאת את לולאת המשוב הפעילה מדי. ניסויים גנטיים מראים שחיזוק או החלשת זרועות שונות במעגל זה יכולים לתקן או להסיר פגמים בהתפשטות ובהורשה, מה שמדגיש כמה המערכת מאוזנת בעדינות.
המשמעויות מעבר לתאי שמרים
הממצאים מראים שביחד, הטרוכרומטין נשמר לא על ידי תגובת "קרא-וכתוב" חלשה אחת, אלא על ידי מעגל משולב של אוביקויטינציה, דה-אצטילציה ומתילציה על חלבוני ההיסטון. בשמרי פיצול, לולאת המשוב H3K14ub–H3K9me3 יוצרת מתג דו-יציב: ברגע שאזור חוצה סף של סימנים אלה, הוא נוטה להישאר שקט ולהתפשט, בעוד אזורים שלא מגיעים לסף לעולם לא מתחייבים במלואם להטרוכרומטין. מאחר ששני סימני ההיסטון הללו והרבה מהאנזימים המעורבים משומרים בממליות, לוגיקה דומה עשויה לסייע לשמור על דפוסי דיכוי גנים יציבים אך גמישים בתאים אנושיים. הבנת המעגל הזה עשויה בסופו של דבר להנחות דרכים חדשות למודולציה של הכרומטין — למשל, לייצוב הגנום בתאים מזדקנים או להעיר באופן סלקטיבי אזורים מושתקים במהלך טיפולי סרטן.
ציטוט: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
מילות מפתח: הטרוכרומטין, שינויי היסטונים, הורשה אפיגנטית, לולאת משוב כרומטין, שמרי פיצול