Clear Sky Science · pl
Obwiednia sprzężenia zwrotnego H3K14ub–H3K9me3 kontroluje rozprzestrzenianie się i dziedziczenie heterochromatyny w drożdżach szczepu fission
Dlaczego ten mikroskopijny przełącznik ma znaczenie
W każdej naszej komórce długie nici DNA są starannie upakowane i zarządzane tak, by właściwe geny włączały się lub wyłączały we właściwym czasie. Główną częścią tego systemu kontroli jest „heterochromatyna” — ściśle upakowane DNA, które utrzymuje pewne odcinki genomu w stanie cichym i stabilnym. Badanie przeprowadzone w drożdżach fission odkrywa ukryty molekularny obwód sprzężenia zwrotnego, który pomaga heterochromatynie rozprzestrzeniać się wzdłuż chromosomów i utrzymywać się podczas podziałów komórkowych. Ponieważ podobne mechanizmy występują u ludzi, zrozumienie tego obwodu daje wskazówki, jak komórki zachowują stabilność genomu i jak jego zaburzenie może przyczyniać się do chorób, a nawet raka.
Ciche sąsiedztwo w genomie
Nie całe DNA jest jednakowo aktywne. Niektóre regiony tworzą heterochromatynę, gdzie geny są w dużej mierze wyłączone. Te ciche odcinki chronią genom, zapobiegając niewłaściwemu zachowaniu sekwencji powtarzalnych, ograniczając szkodliwe rekombinacje oraz kształtując programy ekspresji genów. Heterochromatyna przechodzi trzy kluczowe fazy: inicjuje się w specjalnych miejscach „nukleacji”, rozprzestrzenia do sąsiednich regionów, a następnie jest przekazywana w kolejnych podziałach komórkowych. Chociaż sygnały startowe są stosunkowo dobrze poznane, naukowcy od dawna zastanawiali się, jak stan ciszy wiarygodnie się rozprzestrzenia i utrzymuje, zwłaszcza że wcześniejsze modele opierały się na stosunkowo słabym samowzmacniającym znaczniku na jednym histonie.
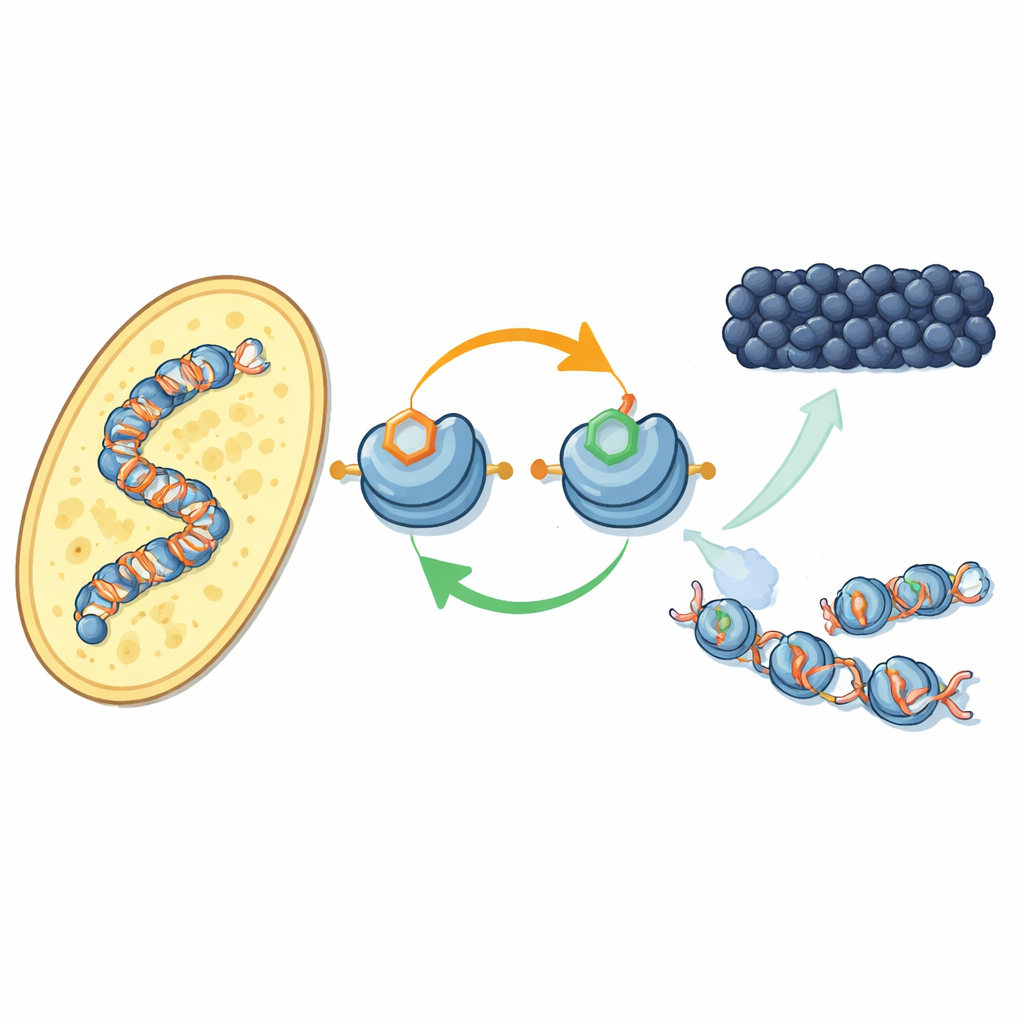
Nowe sprzężenie zwrotne na białkach pakujących DNA
Autorzy koncentrują się na tym, jak konkretne chemiczne znaczniki na histonach — bębnach, wokół których owija się DNA — współdziałają. W drożdżach fission enzym Clr4 dodaje grupę metylową na jedną pozycję w histonie H3 (znaną jako H3K9), co jest cechą heterochromatyny. Ten sam enzym funkcjonuje też w większym kompleksie CLRC, który przyłącza małą grupę ubikwityny na inną pozycję H3 (H3K14). Przy użyciu oczyszczonych składników badacze pokazują, że dołączenie ubikwityny na H3K14 dramatycznie zwiększa zdolność Clr4 do metylacji H3K9 na pełnych nukleosomach, neutralizując w przeciwnym razie hamujący wpływ DNA. Ten efekt stymulujący jest wysoce specyficzny: inne addukcje ubikwityny na różnych pozycjach histonów nie mają takiego samego efektu.
Dwa znaczniki, które wzajemnie się podtrzymują
Przechodząc do żywych komórek drożdży, zespół użył nowo opracowanego przeciwciała, by zmapować występowanie znacznika ubikwityny H3K14 w całym genomie. Występuje on niemal dokładnie tam, gdzie metylacja H3K9 — przy centromerach, telomerach i cichym regionie typu rozmnażania — klasyczne terytoria heterochromatyny. Gdy wyłączyli albo znak metylowy, albo enzymy go umieszczające, znacznik ubikwityny również znikał i odwrotnie. Oba znaczniki są potrzebne, by kompleks CLRC utrzymywał się na chromatynie. Razem ujawnia to dodatnie sprzężenie zwrotne: znacznik ubikwityny wzmacnia metylację, a para znaczników stabilizuje sam kompleks, który je umieszcza, pomagając stanowi ciszy rozprzestrzeniać się po sąsiednich nukleosomach i być przywracanym po replikacji DNA.
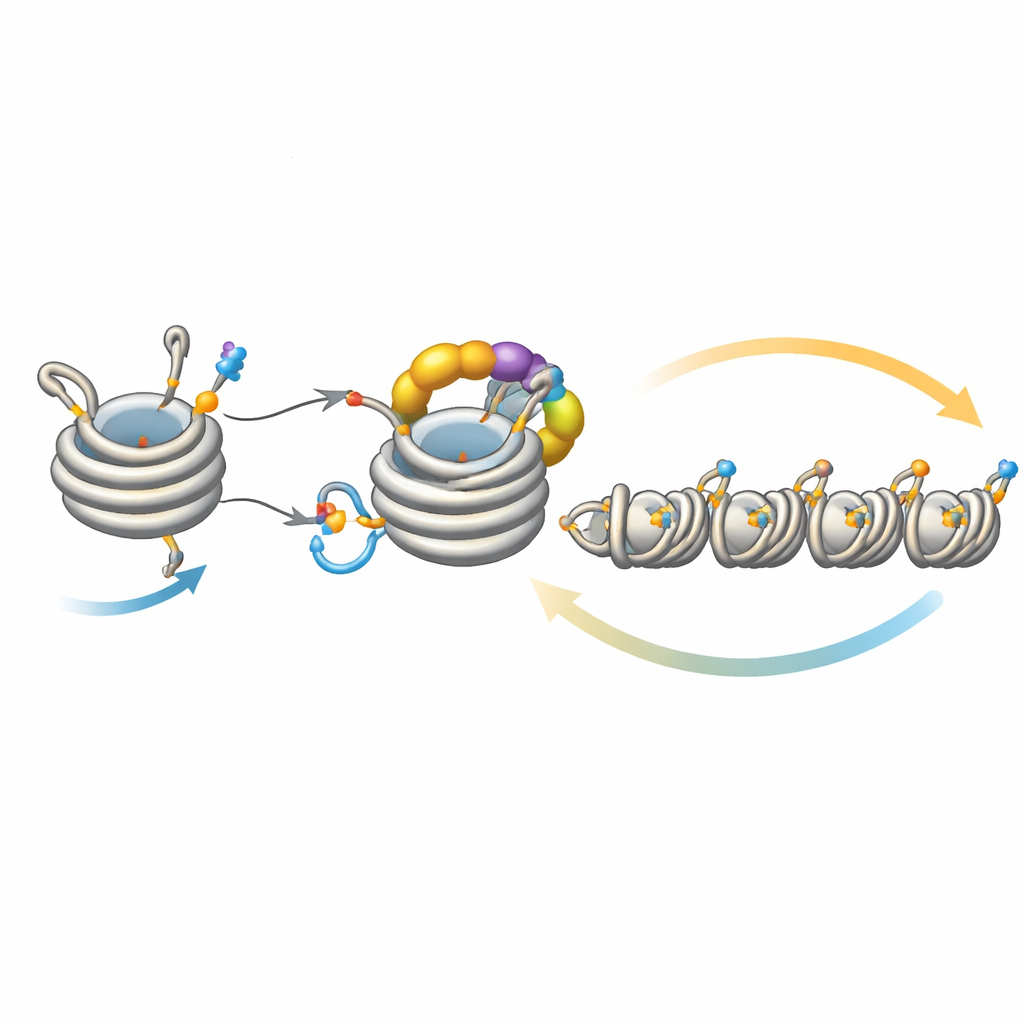
Dostrajać rozprzestrzenianie ciszy
Ten obwód nie działa w izolacji. Deacetylaza Clr3 przygotowuje histon H3 do ubikwitynacji przez usunięcie grupy acetylowej z tego samego miejsca, H3K14; gdy ten krok jest zaburzony, sprzężenie słabnie, a heterochromatyna ma trudności z rozprzestrzenianiem się lub dziedziczeniem, szczególnie w regionach oddalonych od początkowych miejsc nukleacji. Z drugiej strony kilka enzymów działa jak hamulce. Mst2 dodaje grupę acetylową blokującą ubikwitynację, a Epe1 usuwa znacznik metylowy na H3K9. Usunięcie tych hamulców powoduje nadmierne rozprzestrzenianie heterochromatyny, wyciszając geny niezbędne i wywierając stres na komórkach. W takim stresie komórki uruchamiają odpowiedź adaptacyjną, która zmniejsza ilość Clr4, tonując nadaktywne sprzężenie. Eksperymenty genetyczne pokazują, że wzmocnienie lub osłabienie różnych ramion tego obwodu może uratować lub zlikwidować defekty w rozprzestrzenianiu i dziedziczeniu, podkreślając, jak delikatnie wyważony jest ten system.
Implikacje wykraczające poza komórki drożdży
W sumie wyniki pokazują, że heterochromatyna nie jest utrzymywana przez pojedynczą, słabą reakcję „czytaj-pisz”, lecz przez zintegrowany układ ubikwitynacji, deacetylacji i metylacji na białkach histonowych. W drożdżach fission pętla sprzężenia H3K14ub–H3K9me3 tworzy bistabilny przełącznik: gdy region przekroczy próg tych znaczników, ma skłonność pozostawać cichy i się rozprzestrzeniać, podczas gdy regiony, które tego progu nie osiągają, nigdy w pełni nie przechodzą w heterochromatynę. Ponieważ oba te modyfikacje histonów i wiele zaangażowanych enzymów są zachowane u ssaków, podobna logika może pomagać utrzymywać stabilne, a zarazem elastyczne wzorce represji genów w komórkach ludzkich. Zrozumienie tej sieci może ostatecznie pomóc w opracowaniu nowych sposobów modulacji chromatyny — na przykład w celu stabilizacji genomu w komórkach starzejących się lub selektywnego przywracania aktywności cichych regionów podczas terapii przeciwnowotworowej.
Cytowanie: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
Słowa kluczowe: heterochromatyna, modyfikacja histonów, dziedziczenie epigenetyczne, sprzężenie zwrotne chromatyny, drożdże fission