Clear Sky Science · es
Un circuito de retroalimentación H3K14ub-H3K9me3 gobierna la propagación y la herencia de la heterocromatina en la levadura fisionante
Por qué importa este interruptor microscópico
Dentro de cada una de nuestras células, largas hebras de ADN se empaquetan y gestionan con cuidado para que los genes adecuados se activen o desactiven en el momento correcto. Una parte importante de este sistema de control es la «heterocromatina», ADN densamente empaquetado que mantiene silenciosos y estables ciertos tramos del genoma. Este estudio, realizado en la levadura fisionante, descubre un circuito molecular de retroalimentación oculto que ayuda a que la heterocromatina se propague a lo largo de los cromosomas y persista cuando las células se dividen. Dado que existen sistemas similares en humanos, comprender este circuito ofrece pistas sobre cómo las células mantienen la estabilidad genómica y cómo su fallo puede contribuir a enfermedades e incluso al cáncer.
Un vecindario silencioso en el genoma
No todo el ADN es igualmente activo. Algunas regiones forman heterocromatina, donde los genes están mayoritariamente apagados. Estos tramos silenciosos protegen el genoma al impedir que el ADN repetitivo se comporte de forma errática, limitar recombinaciones dañinas y configurar programas de expresión génica. La heterocromatina tiene tres fases clave: se inicia en sitios especiales de «nucleación», se propaga a las regiones vecinas y luego se transmite a través de las divisiones celulares. Aunque las señales de inicio están bastante bien entendidas, los científicos llevan tiempo intrigados por cómo el estado silencioso se propaga y perdura de forma fiable, especialmente dado que los modelos previos dependían de una marca auto-reforzante relativamente débil en una histona.
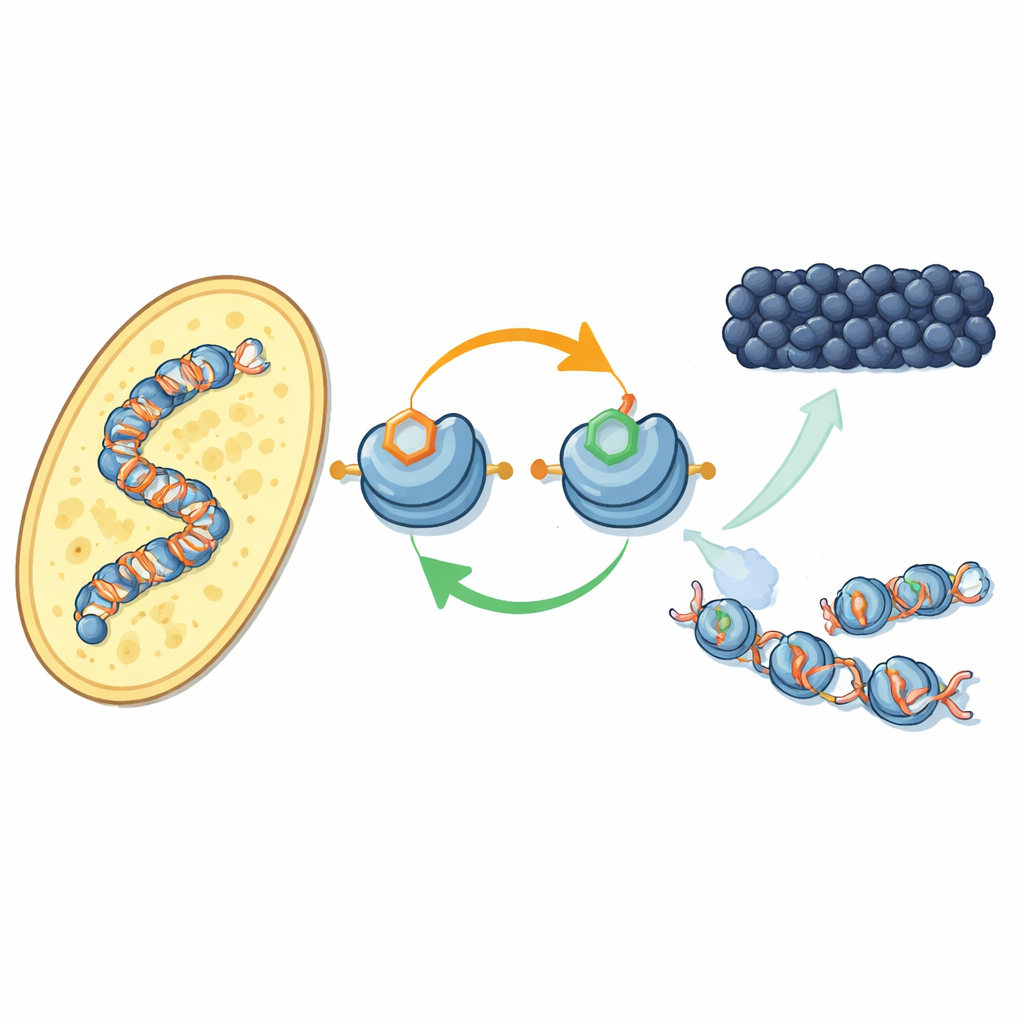
Un nuevo bucle de retroalimentación sobre proteínas que empaquetan el ADN
Los autores se centran en cómo etiquetas químicas específicas en las histonas —los carretes alrededor de los que se enrolla el ADN— actúan en conjunto. En la levadura fisionante, una enzima llamada Clr4 añade una marca de metilo en una posición de la histona H3 (conocida como H3K9), sello característico de la heterocromatina. La misma enzima también forma parte de un complejo mayor, CLRC, que coloca una pequeña etiqueta de ubiquitina en otra posición de H3 (H3K14). Usando componentes purificados, los investigadores muestran que añadir ubiquitina en H3K14 aumenta de forma dramática la capacidad de Clr4 para metilar H3K9 en nucleosomas completos, superando un efecto inhibitor del ADN. Este efecto estimulador es muy específico: otras ubiquitinaciones en posiciones distintas de las histonas no tienen el mismo impacto.
Dos marcas que se sostienen mutuamente
En células vivas de levadura, el equipo empleó un anticuerpo recién desarrollado para mapear dónde aparece la etiqueta de ubiquitina en H3K14 a lo largo del genoma. Aparece casi exactamente donde está la metilación de H3K9: en centrómeros, telómeros y una región silente de tipo de apareamiento —territorios clásicos de heterocromatina. Cuando desactivaron la marca de metilo o las enzimas que la colocan, la marca de ubiquitina también desapareció, y viceversa. Ambas marcas son necesarias para mantener anclado al complejo CLRC sobre la cromatina. En conjunto, esto revela un bucle de retroalimentación positivo: la ubiquitina potencia la metilación, y el par de marcas estabiliza el propio complejo que las coloca, ayudando a que el estado silencioso se propague a nucleosomas vecinos y se restablezca después de la replicación del ADN.
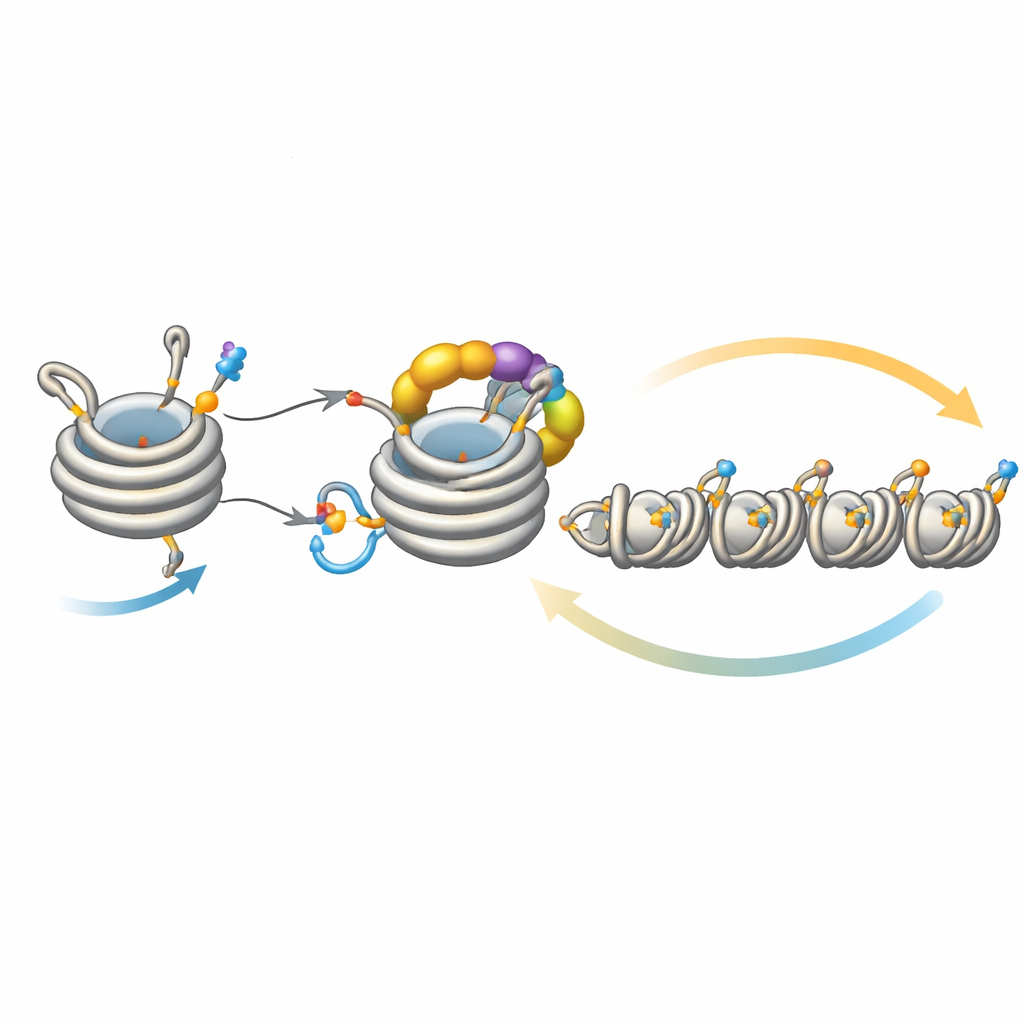
Afinar la extensión del silencio
Este bucle no funciona en aislamiento. Una desacetilasa llamada Clr3 prepara la histona H3 para la ubiquitinación al eliminar un grupo acetilo en el mismo sitio, H3K14; cuando este paso se ve comprometido, la retroalimentación se debilita y la heterocromatina tiene dificultades para propagarse o heredarse, especialmente en regiones alejadas de los sitios de nucleación originales. Por el contrario, varias enzimas actúan como frenos. Mst2 añade el grupo acetilo que bloquea la ubiquitinación, y Epe1 elimina la marca de metilo en H3K9. Quitar estos frenos provoca que la heterocromatina se propague en exceso, silenciando genes esenciales y estresando a las células. Bajo tal estrés, las células disparan una respuesta adaptativa que reduce la cantidad de Clr4, moderando el bucle de retroalimentación sobreactivado. Experimentos genéticos muestran que reforzar o debilitar distintos brazos de este circuito puede rescatar o abolir defectos en la propagación y la herencia, lo que subraya lo delicadamente equilibrado que está el sistema.
Implicaciones más allá de las células de levadura
En conjunto, los hallazgos revelan que la heterocromatina no se mantiene por una sola reacción «leer-escribir» débil, sino por un circuito integrado de ubiquitinación, desacetilación y metilación en proteínas histónicas. En la levadura fisionante, este bucle H3K14ub–H3K9me3 crea un interruptor bistable: una vez que una región supera un umbral de estas marcas, tiende a permanecer silenciosa y a propagarse, mientras que las regiones que no lo hacen nunca llegan a comprometerse plenamente con la heterocromatina. Dado que ambas marcas histónicas y muchas de las enzimas implicadas se conservan en mamíferos, una lógica similar podría ayudar a mantener patrones de represión génica estables pero flexibles en células humanas. Comprender esta circuitería podría eventualmente informar nuevas formas de modular la cromatina —por ejemplo, para estabilizar el genoma en células envejecidas o para reactivar selectivamente regiones silenciosas durante terapias contra el cáncer.
Cita: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
Palabras clave: heterocromatina, modificación de histonas, herencia epigenética, bucle de retroalimentación de la cromatina, levadura fisionante