Clear Sky Science · ru
Контур обратной связи H3K14ub–H3K9me3 управляет распространением и наследованием гетерохроматина у дробянки
Почему этот микроскопический переключатель важен
Внутри каждой нашей клетки длинные молекулы ДНК аккуратно упакованы и управляются так, чтобы нужные гены включались или выключались в нужное время. Важной частью этой системы контроля является «гетерохроматин» — плотно упакованная ДНК, которая удерживает определённые участки генома в молчании и стабильности. В этом исследовании на дробянке обнаружен скрытый молекулярный контур обратной связи, который помогает гетерохроматину распространяться вдоль хромосом и сохраняться при делении клеток. Поскольку похожие механизмы существуют у человека, понимание этого контура даёт подсказки о том, как клетки поддерживают стабильность генома и как нарушение этого механизма может способствовать заболеваниям и даже раку.
Тихий квартал в геноме
Не вся ДНК активна в равной степени. Некоторые участки формируют гетерохроматин, где гены по большей части выключены. Эти молчаливые зоны защищают геном, предотвращая ненадлежащее поведение повторяющихся последовательностей, ограничивая вредные рекомбинации и формируя программы экспрессии генов. У гетерохроматина три ключевых стадии: он начинается на специальных «нуклеационных» участках, распространяется на соседние регионы и затем передаётся при делениях клеток. В то время как стартовые сигналы в целом понятны, учёные долго недоумевали, как состояние молчания надёжно распространяется и сохраняется, особенно учитывая, что прежние модели опирались на относительно слабую самоподдерживающую метку на одном белке-гистоне.
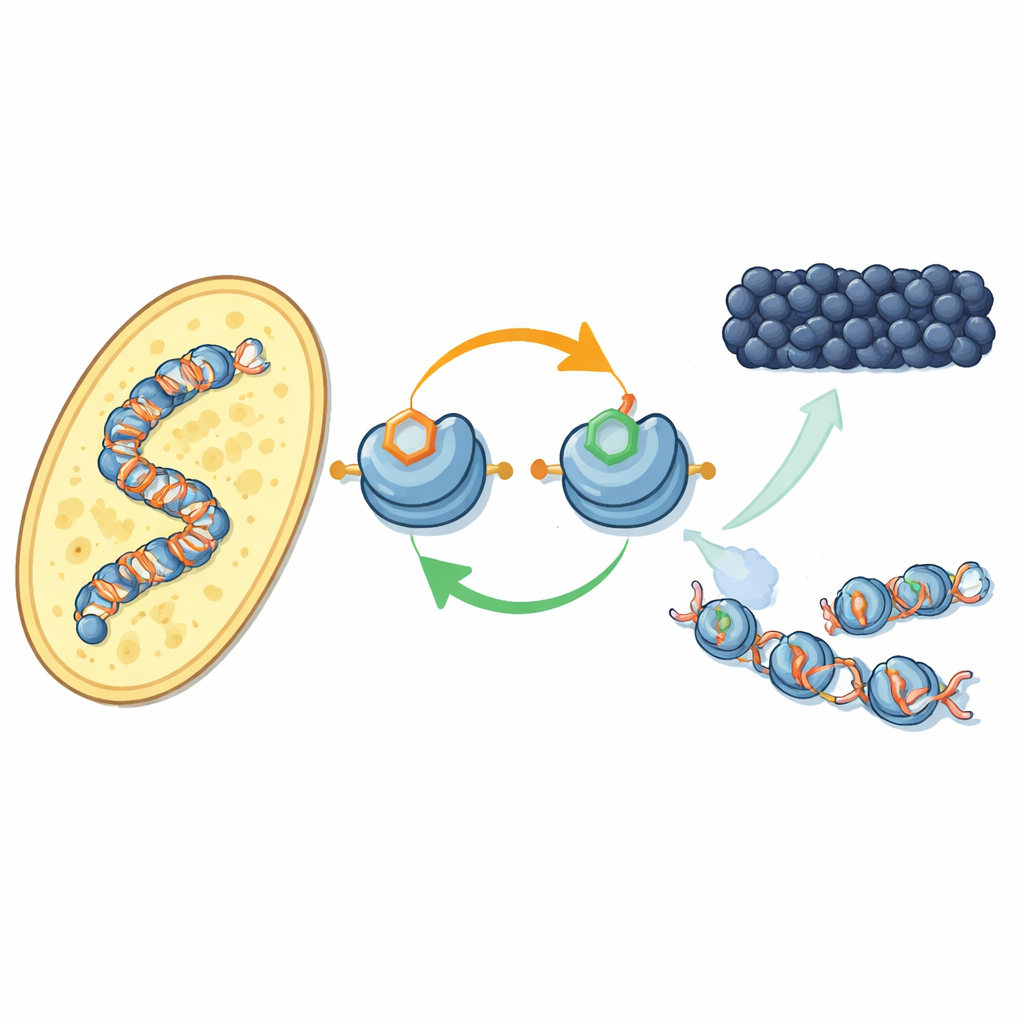
Новый контур обратной связи на белках упаковки ДНК
Авторы сосредоточились на том, как специфические химические метки на белках-гистонах — шпульках, вокруг которых намотана ДНК — взаимодействуют. У дробянки фермент Clr4 добавляет метильную метку в одну позицию на гистоне H3 (известную как H3K9), что является характерной чертой гетерохроматина. Тот же фермент входит в больший комплекс CLRC, который прикрепляет небольшой убиквитин на другую позицию H3 (H3K14). На очищенных компонентах исследователи показали, что убиквитинирование H3K14 резко усиливает способность Clr4 метилировать H3K9 на полных нуклеосомах, преодолевая в противном случае ингибирующее влияние ДНК. Этот стимулирующий эффект очень специфичен: другие убиквитиновые метки на разных позициях гистонов не оказывают такого же влияния.
Две метки, которые поддерживают друг друга
Перейдя к живым клеткам дробянки, команда использовала недавно созданный антитело, чтобы отобразить, где в геноме появляется убиквитиновая метка на H3K14. Она находится почти в тех же местах, что и метилирование H3K9 — в центромерах, теломерах и в молчаливом участке, ответственном за тип спаривания — классические территории гетерохроматина. Когда они отключали либо метильную метку, либо ферменты, её устанавливающие, убиквитиновая метка также исчезала, и наоборот. Обе метки необходимы, чтобы комплекс CLRC удерживался на хроматине. В совокупности это раскрывает положительный контур обратной связи: убиквитин усиливает метилирование, а пара меток стабилизирует сам комплекс, который их ставит, помогая молчаливому состоянию распространяться на соседние нуклеосомы и восстанавливаться после репликации ДНК.
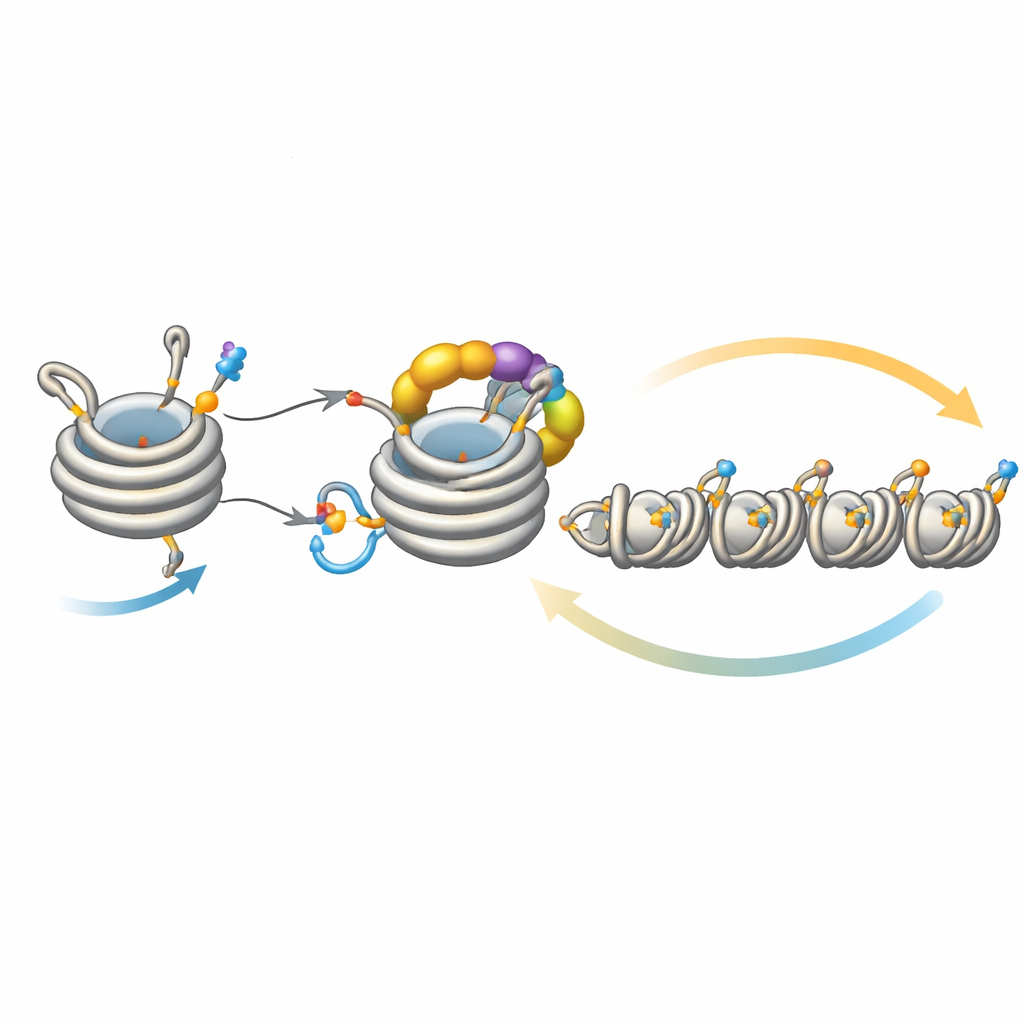
Тонкая настройка распространения молчания
Этот контур не действует в изоляции. Деацетилаза Clr3 подготавливает гистон H3 к убиквитинированию, удаляя ацетильную группу с той же позиции H3K14; при нарушении этого шага контур ослабевает, и гетерохроматин с трудом распространяется или наследуется, особенно в регионах, удалённых от исходных нуклеационных участков. Напротив, несколько ферментов действуют как тормоза. Mst2 добавляет ацетильную группу, блокирующую убиквитинирование, а Epe1 удаляет метильную метку с H3K9. Удаление этих тормозов заставляет гетерохроматин распространяться слишком далеко, и важные гены отключаются, что вызывает стресс в клетках. В ответ на такой стресс клетки запускают адаптивную реакцию, уменьшающую количество Clr4 и ослабляющую чрезмерно активный контур обратной связи. Генетические эксперименты показывают, что усиление или ослабление разных звеньев этого контура может восстановить или уничтожить способность к распространению и наследованию, подчёркивая, насколько тонко сбалансирована система.
Значение за пределами дробянки
Вместе эти результаты показывают, что гетерохроматин поддерживается не одной слабой «читай-пиши» реакцией, а интегрированным контуром убиквитинирования, деацетилирования и метилирования на белках-гистонах. У дробянки контур H3K14ub–H3K9me3 создаёт би-стабильный переключатель: как только регион превышает порог этих меток, он склонен оставаться молчаливым и распространяться, тогда как регионы, не достигшие порога, так и не переходят в гетерохроматин. Поскольку обе эти метки гистонов и многие вовлечённые ферменты консервативны у млекопитающих, аналогичная логика может помогать поддерживать стабильные, но гибкие шаблоны репрессии генов в клетках человека. Понимание этой схемы в перспективе может подсказать новые способы модуляции хроматина — например, для стабилизации генома в стареющих клетках или для избирательного пробуждения молчаливых участков при терапии рака.
Цитирование: Toda, T., Zang, J., Qi, H. et al. An H3K14ub-H3K9me3 feedback circuit governs heterochromatin spreading and inheritance in fission yeast. Nat Commun 17, 3483 (2026). https://doi.org/10.1038/s41467-026-70276-8
Ключевые слова: гетерохроматин, модификация гистонов, эпигенетическое наследование, контур обратной связи хроматина, дробянка