Clear Sky Science · tr
Effektor konformasyonel plastisitesi, bağırsak simbiontlerinde Hcp heterohekzamerleri aracılığıyla soy-spesifik salgılamayı mümkün kılar
İnsan bağırsağında mikrobiyal düellolar
Bağırsaklarımız, sürekli olarak yer ve besin için itişip duran yoğun mikrop topluluklarına ev sahipliği yapar. Hayatta kalmak için bazı bakteriler, rakiplerini delip zehirleyebilen moleküler silahlar kullanır; bu da hangi türlerin bağırsakta gelişeceğini incelikle şekillendirir. Bu çalışma, yaygın bir insan bağırsak simbiontu olan Bacteroides fragilis’in, belirli toksik proteinleri komşu bakterilere olağanüstü bir kesinlikle teslim edebilmesi için küçük bir enjeksiyon aparatı nasıl özelleştirdiğini ortaya koyuyor.
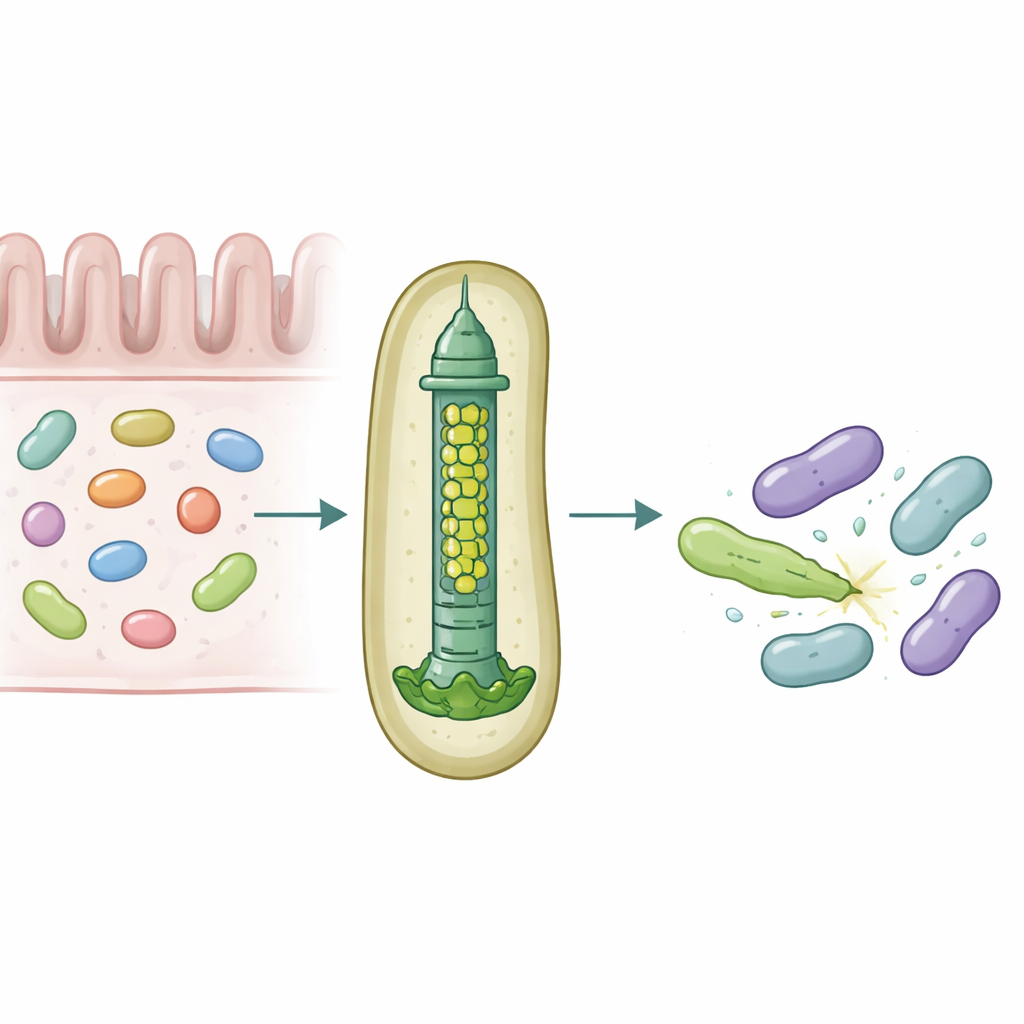
Bakteriyel savaş için mikroskobik bir mızrak tabancası
Çok sayıda Gram-negatif bakteri, ters çevrilmiş bir faj kuyruğu gibi çalışan yay yüklü bir yapı olan Tip VI Salgılama Sistemi adlı bir aygıtı kullanarak toksik “effektor”leri yakın hücrelere fırlatır. Bu sistemin merkezi bir parçası, Hcp adı verilen bir proteinden yapılmış üst üste dizilmiş halkalardan oluşan bir borudur. Çoğu türde her sistemin tek bir Hcp formu bulunurken, insan bağırsağından izole edilen B. fragilis aynı lokusta beş farklı hcp genine sahiptir. Bu görünen fazlalığın biyolojik nedeni bilinmiyordu; özellikle de bu sistemin laboratuvar ve hayvan modellerinde yakın akraba bakterileri öldürmede olağanüstü derecede etkili olması nedeniyle.
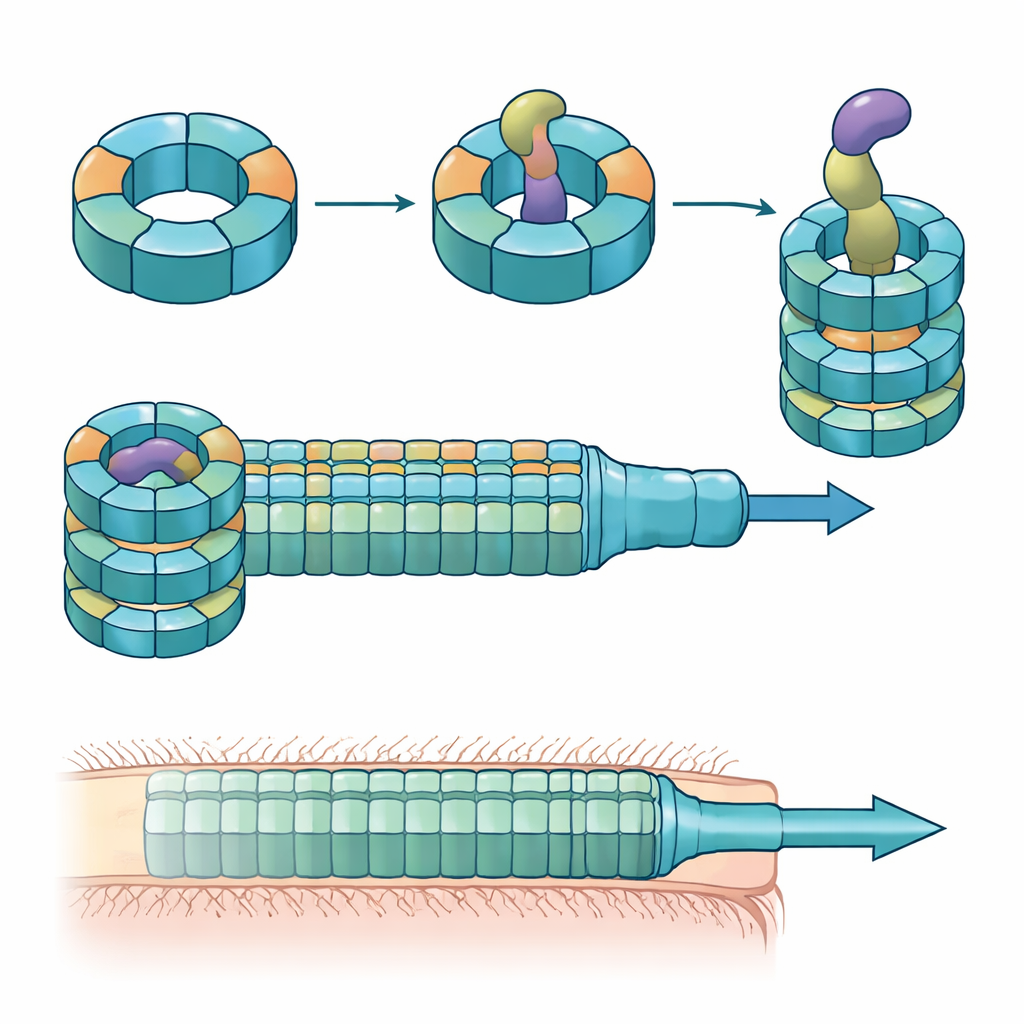
Karışık parçalarla inşa edilmiş uzmanlaşmış halkalar
Araştırmacılar, fare kolonizasyon deneyleri, karışık bakteri kültürleri ve protein biyokimyası yöntemlerini birleştirerek bu Hcp varyantlarının rollerini çözdüler. Belirli bir effektor olan Bte1 tarafından etkin öldürme için yalnızca Hcp1, Hcp2 ve Hcp3’ün gerekli olduğunu buldular. Beklendiği üzere Hcp1 salgılama makinesinin ana tüpünü oluşturuyor. Ancak Hcp2 ve Hcp3 farklı bir şey yapıyor: bunlar birlikte, ya tek başlarına oluşturulamayan altı parçalı karışık bir halka, bir heterohekzamer halinde monte oluyorlar. Kriyomikro-elektron mikroskobu kullanarak ekip, bu heterohekzamerlerin simetrik bir halka içinde düzenlenmiş dört Hcp2 ve iki Hcp3 ünitesinden oluştuğunu; sert bir gözenek yarattığını, Bte1 ile etkileşebildiğini fakat Hcp1 halkalarıyla karışmadığını gösterdi.
Şekil değiştiren yük ve kilit-anahtar uyumu
Hcp2–Hcp3 halkasının Bte1’e bağlı yüksek çözünürlüklü yapıları, effektörün merkezi kanala sıkıca oturabilmek için şeklinin bazı bölümlerini dramatik şekilde yeniden düzenlemesi gerektiğini ortaya koydu. Çeşitli α-heliksleri, protein tüple çakışmayı önlemek için onlarla on derece mertebesinde açısal kaymalar gösteriyordu; yazarların konformasyonel plastisite dediği şey bu. Bu helikslerden birini sabitleyecek bir disülfit bağı mühendisliklediklerinde, Bte1 artık halkaya etkin bağlanamadı ve rekabet deneylerinde öldürme etkinliği düştü. Temas yüzeylerinin ayrıntılı haritalanması, bağlanma ve salgılama için hem Hcp2 hem de Hcp3 üzerindeki belirli amino asitlerin gerekli olduğunu ortaya koydu. Sadece bir kalıntının karşı tarafta değiştirilmesi, halkayı kendisi parçalamadan teslimatı bozmak için yeterliydi.
Bağırsak soyları arasında birlikte evrimleşen halkalar ve toksinler
Takım daha sonra bu mekanizmanın tek bir suşa özgü olup olmadığını yoksa B. fragilis soyları arasında paylaşılıp paylaşılmadığını sordu. Onlarca suştan Hcp2–Hcp3 çiftlerini ve yakınlardaki effektor genlerini karşılaştırarak, her soyun lokusun değişken bir bölgesinde karakteristik bir effektöre ve eşleşen bir Hcp2–Hcp3 çiftine sahip olduğunu buldular. Bu çiftleri suşlar arasında çapraz değiş tokuş etmek, her karışık halkanın kendi effektörünü tanıdığını gösterdi; farklı halkalar yapısal olarak hâlâ bir araya gelebilse bile. Özellikle Hcp3, halkanın iç yüzeyinde yüksek değişkenlik gösteren yamalar içeriyor ve hangi effektörün bağlanabileceğinin ana belirleyicisi gibi davranıyor. Bu dizi çeşitliliğine rağmen, effektörlerin kendileri yeniden kullanılabilir bir modül gibi halkaya yerleşen korunan bir N-terminal katlanma paydasını paylaşıyor; C-terminal toksik bölgeleri ise büyük ölçüde farklılık gösteriyor.
Bu şekil-eşleme sisteminin önemi
Bu bulgular birlikte bir “kilit-ve-anahtar” eşevrimsel modelini tasvir ediyor: Hcp2–Hcp3 halkasının iç yüzeyi soy-spesifik bir kilit sağlar ve her effektörün korunan N-terminal modülü farklı suşlarda ince şekilde yeniden şekillendirilmiş bir anahtar işlevi görür. Effektör proteinler, boruyu tıkamadan halkanın içinden geçecek kadar esneyip yeniden katlanabilir; bu, nispeten büyük veya karmaşık toksinlerin dar bir kanaldan teslim edilmesine izin verir. Bu çalışma, bağırsak simbiontlerinin moleküler mızraklarını nasıl ince ayar yaptıklarını ve mikrobiyal rakiplere karşı farklı toksinler kullanabildiklerini açıklıyor; ayrıca bu kilit-ve-anahtar arayüzlerini yeniden kablolarak, araştırmacıların bir gün Tip VI sistemlerini mikrobiyal topluluklarda yararlı moleküller için programlanabilir teslimat platformlarına dönüştürebileceğini öne sürüyor.
Atıf: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Anahtar kelimeler: Tip VI salgılama sistemi, Bacteroides fragilis, bakteriyel rekabet, protein konformasyonel plastisitesi, mikrobiyom mühendisliği