Clear Sky Science · de
Konformationelle Plastizität von Effektoren ermöglicht linien-spezifische Sekretion über Hcp‑Heterohexamere bei Darmsymbionten
Mikrobielle Duelle im menschlichen Darm
In unserem Darm leben dichte Mikrobengemeinschaften, die ständig um Raum und Nährstoffe konkurrieren. Zum Überleben setzen manche Bakterien molekulare Waffen ein, die Rivalen durchstechen und vergiften können und so dezent bestimmen, welche Arten im Darm gedeihen. Diese Studie enthüllt, wie ein häufiger menschlicher Darmsymbiont, Bacteroides fragilis, ein winziges Injektionsgerät maßschneidert, damit es bestimmte toxische Proteine mit bemerkenswerter Präzision in benachbarte Bakterien abgeben kann.
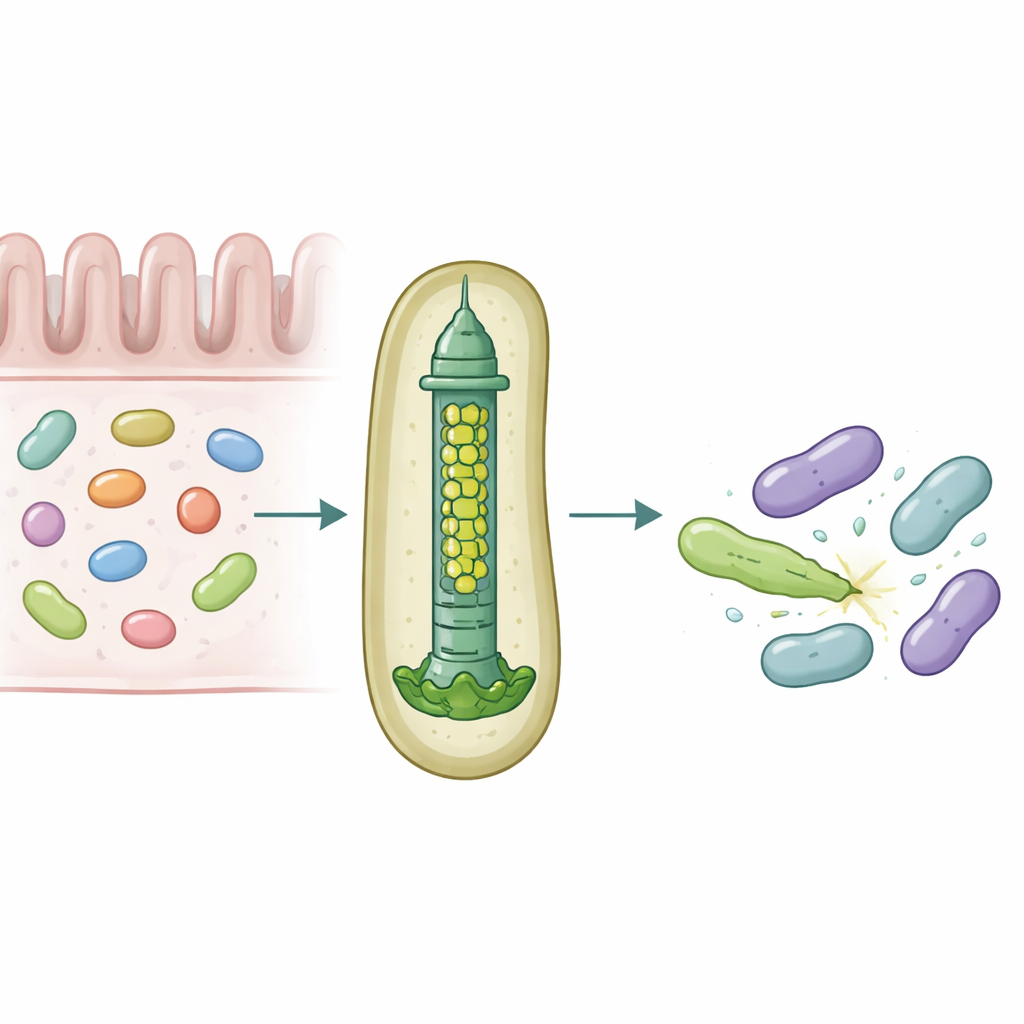
Eine mikroskopische Speerschleuder für bakterielle Kriegsführung
Viele gramnegative Bakterien nutzen ein Gerät namens Type‑VI‑Sekretionssystem, eine federbelastete Struktur, die wie ein umgekehrter Phagen‑Schwanz funktioniert, um toxische „Effektoren“ in nahegelegene Zellen zu schleudern. Ein zentraler Teil dieses Systems ist ein Rohr, aufgebaut aus übereinandergestapelten Ringen eines Proteins namens Hcp. Bei den meisten Arten gibt es pro System nur eine Form von Hcp, doch B. fragilis aus dem menschlichen Darm trägt fünf verschiedene hcp‑Gene in einem einzigen Locus. Der biologische Grund für diese scheinbare Redundanz war unklar, zumal dieses System in Labor‑ und Tiermodellen ungewöhnlich wirksam beim Töten eng verwandter Bakterien ist.
Spezialisierte Ringe aus gemischten Bauteilen
Die Forschenden kombinierten Kolonisierungsexperimente in Mäusen, gemischte Bakterienkulturen und Proteinbiochemie, um die Rolle dieser Hcp‑Varianten zu analysieren. Sie fanden heraus, dass nur drei — Hcp1, Hcp2 und Hcp3 — für ein effizientes Töten durch einen bestimmten Effektor namens Bte1 erforderlich sind. Hcp1 bildet wie erwartet das Hauptrohr der Sekretionsmaschine. Hcp2 und Hcp3 hingegen tun etwas anderes: sie setzen sich gemeinsam zu einem gemischten sechs‑teiligen Ring zusammen, einem Heterohexamer, das aus keinem der beiden Proteine allein gebildet werden kann. Mithilfe von Kryo‑Elektronenmikroskopie zeigte das Team, dass diese Heterohexamere aus vier Hcp2‑ und zwei Hcp3‑Einheiten bestehen, die zu einem symmetrischen Ring angeordnet sind und eine starre Pore bilden, die Bte1 binden kann, sich aber nicht mit Hcp1‑Ringen vermischt.
Formwandelnder Frachtstoff und ein Schloss‑und‑Schlüssel‑Prinzip
Hochaufgelöste Strukturen des Hcp2–Hcp3‑Rings gebunden an Bte1 zeigten, dass der Effektor Teile seiner Gestalt dramatisch umordnen muss, um sich eng in den zentralen Kanal einzufügen. Mehrere seiner α‑Helices schwenken um Zehnergrade, um Kollisionen mit dem Proteintubus zu vermeiden — ein Beispiel für die von den Autor:innen so bezeichnete konformationelle Plastizität. Als sie eine Disulfidbrücke einbauten, um eine dieser Helices zu fixieren, konnte Bte1 nicht mehr effizient an den Ring binden, und seine Toxizität fiel in Konkurrenzexperimenten ab. Detaillierte Kartierungen der Kontaktflächen identifizierten spezifische Aminosäuren sowohl auf Hcp2 als auch auf Hcp3, die für Bindung und Sekretion essentiell sind. Schon die Mutation einer einzigen Restgruppe auf einer der Partner reichte aus, um die Lieferung zu stören, ohne den Ring selbst zu zerstören.
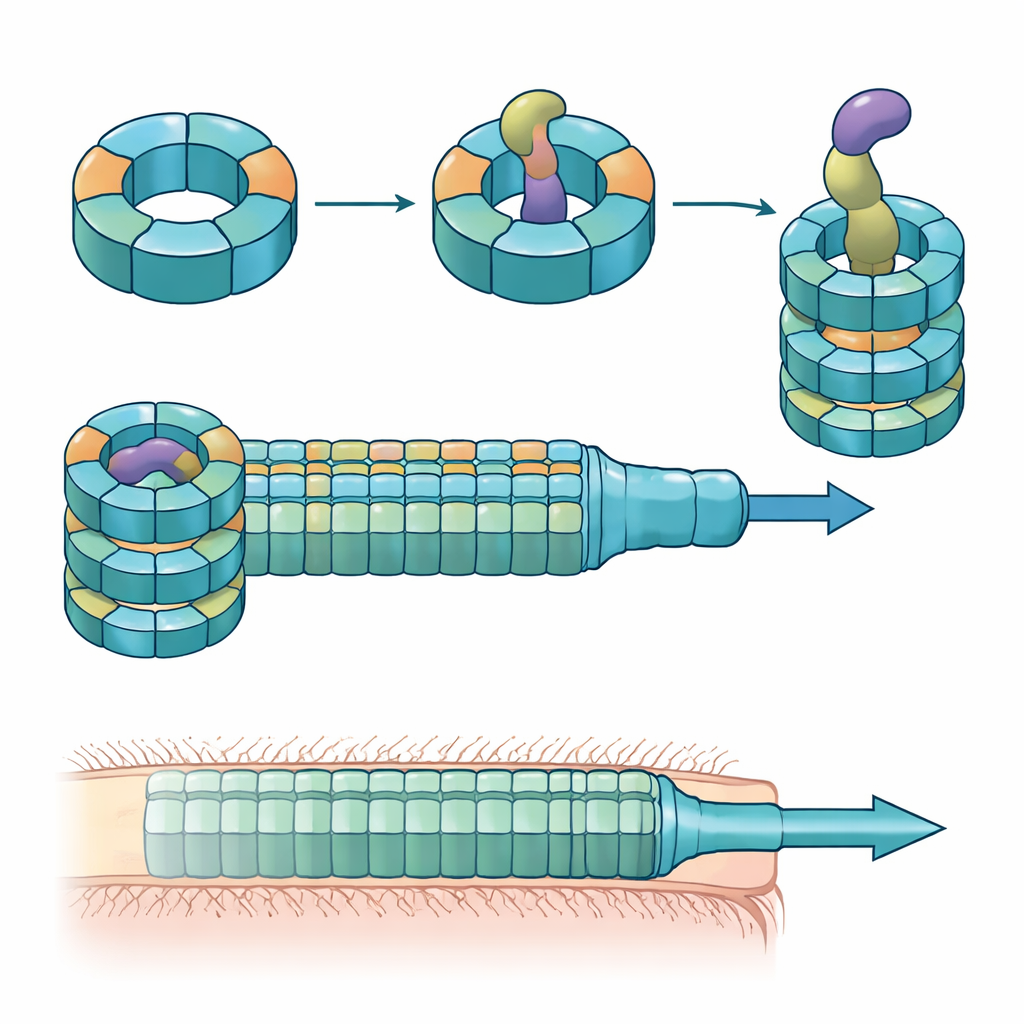
Kojen‑weise mitfunktionierende Ringe und Toxine über Darmlinien hinweg
Das Team untersuchte anschließend, ob dieser Mechanismus auf einen einzelnen Stamm beschränkt ist oder in verschiedenen B. fragilis‑Linien vorkommt. Beim Vergleich von Hcp2–Hcp3‑Paaren und ihren benachbarten Effektorgenen aus Dutzenden von Stämmen stellten sie fest, dass jede Linie in einer variablen Region des Locus einen charakteristischen Effektor und ein dazu passendes Hcp2–Hcp3‑Paar trägt. Das Kreuztauschen dieser Paare zwischen Stämmen zeigte, dass jeder gemischte Ring nur seinen eigenen Effektor erkennt, obwohl verschiedene Ringe strukturell weiterhin zusammengebaut werden können. Insbesondere Hcp3 enthält stark variable Flächen auf der Innenoberfläche des Rings, die als Hauptbestimmungsfaktor dafür dienen, welcher Effektor binden kann. Trotz dieser Sequenzvielfalt teilen die Effektoren selbst eine konservierte N‑terminale Faltung, die wie ein wiederverwendbares Modul in den Ring einrastet, während ihre C‑terminalen toxischen Domänen stark variieren.
Warum dieses Form‑Matching‑System wichtig ist
Diese Ergebnisse skizzieren zusammen ein Ko‑evolutionsmodell im Sinne eines „Schloss‑und‑Schlüssel“‑Prinzips: Die Innenfläche des Hcp2–Hcp3‑Rings bildet ein linien‑spezifisches Schloss, und das konservierte N‑terminale Modul jedes Effektors dient als Schlüssel, der in verschiedenen Stämmen subtil umgestaltet wurde. Effektorproteine können sich flexibel anpassen und gerade genug umfalten, um durch den Ring zu passen, ohne das Rohr zu blockieren, sodass relativ große oder komplexe Toxine durch einen schmalen Kanal geliefert werden können. Diese Arbeit erklärt, wie Darmsymbionten ihre molekularen Speerschleudern feinabstimmen, um unterschiedliche Toxine gegen mikrobielle Rivalen einzusetzen, und sie legt nahe, dass man durch das Umlenken dieser Schloss‑und‑Schlüssel‑Schnittstellen eines Tages Type‑VI‑Systeme als programmierbare Lieferplattformen für nützliche Moleküle in mikrobiellen Gemeinschaften konstruieren könnte.
Zitation: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Schlüsselwörter: Type‑VI‑Sekretionssystem, Bacteroides fragilis, Bakterienkonkurrenz, Protein‑Konformationsplastizität, Microbiom‑Engineering