Clear Sky Science · it
La plasticità conformazionale degli effettori consente secrezione specifica per lignaggio tramite eteroesameri Hcp nei simbionti intestinali
Duelli microbici all'interno dell'intestino umano
I nostri intestini ospitano comunità dense di microbi che si contendono continuamente spazio e nutrimento. Per sopravvivere, alcuni batteri impiegano armi molecolari in grado di perforare e avvelenare i rivali, influenzando in modo sottile quali specie prosperano nell'intestino. Questo studio svela come un comune simbionte intestinale umano, Bacteroides fragilis, personalizzi un minuscolo apparato in grado di iniettare tossine specifiche nelle cellule batteriche vicine con notevole precisione.
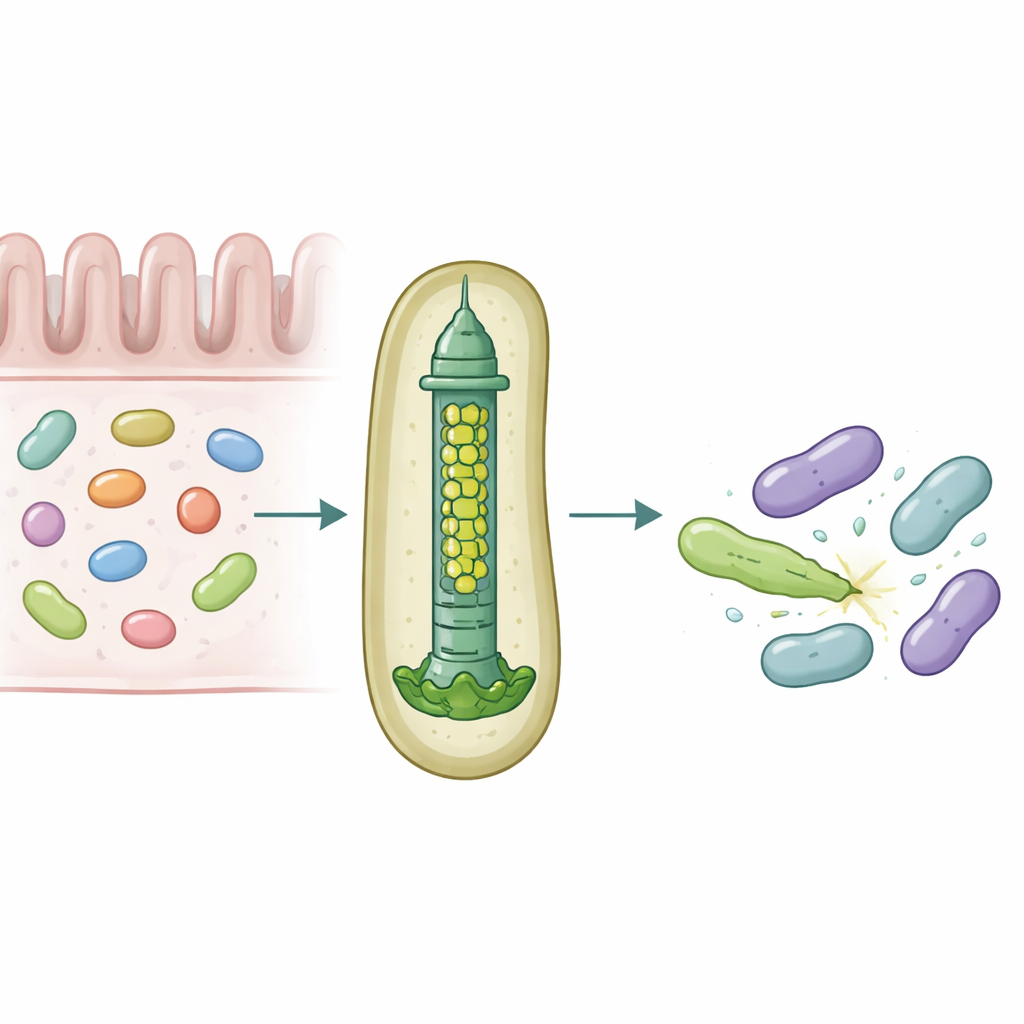
Una microlancia per la guerra batterica
Molti batteri Gram-negativi usano un dispositivo chiamato Sistema di Secrezione di Tipo VI, una struttura carica a molla che funziona come una coda di fago capovolta, per sparare «effettori» tossici nelle cellule vicine. Una componente centrale di questo sistema è un tubo formato da anelli impilati di una proteina chiamata Hcp. Nella maggior parte delle specie ogni sistema possiede una sola variante di Hcp, mentre B. fragilis dell'intestino umano porta cinque diversi geni hcp in un unico locus. Il motivo biologico di questa apparente ridondanza era sconosciuto, soprattutto perché questo sistema è straordinariamente efficiente nell'uccidere batteri strettamente imparentati in modelli di laboratorio e animali.
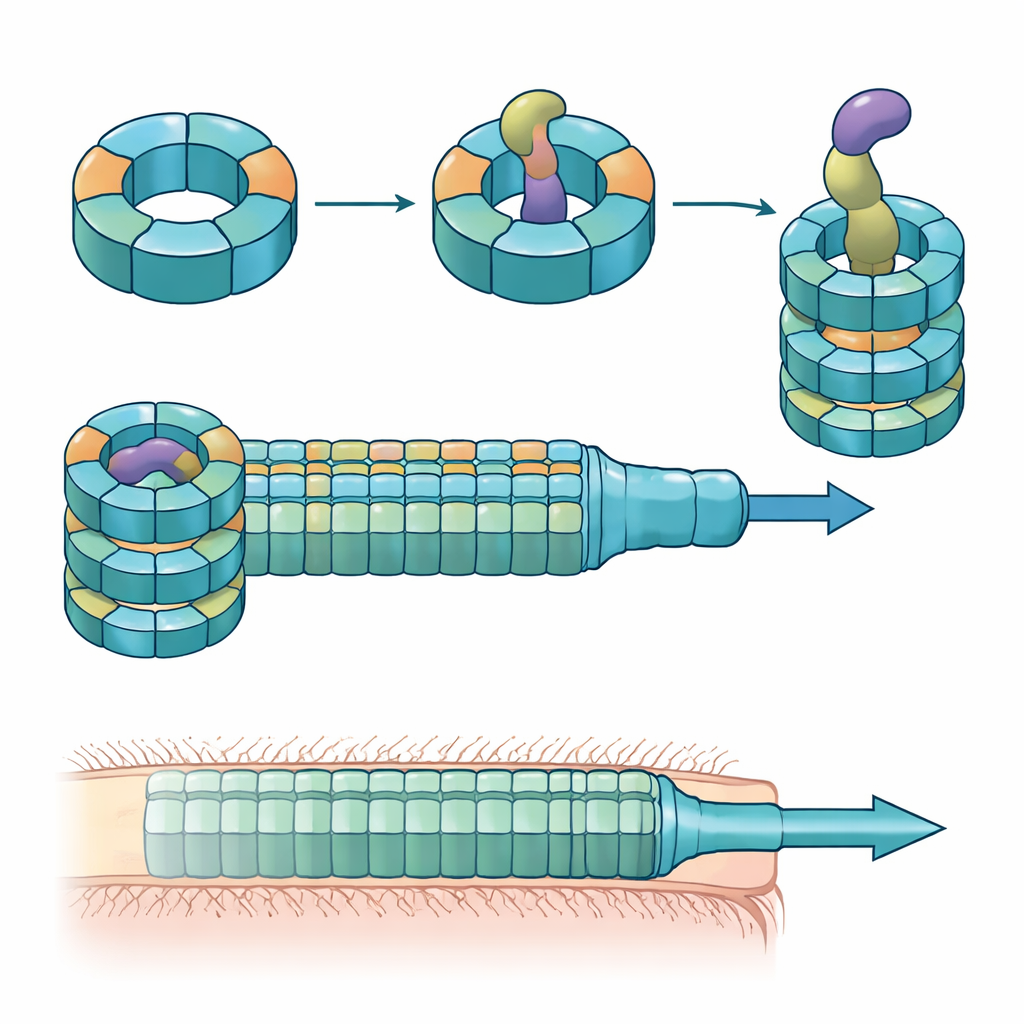
Anelli specializzati costruiti con parti miste
Gli autori hanno combinato esperimenti di colonizzazione in topo, colture batteriche miste e biochimica delle proteine per disassemblare i ruoli di queste varianti di Hcp. Hanno scoperto che solo tre—Hcp1, Hcp2 e Hcp3—sono necessari per un'uccisione efficiente da parte di un effettore chiamato Bte1. Hcp1 forma il tubo principale della macchina di secrezione, come previsto. Hcp2 e Hcp3, invece, fanno qualcosa di diverso: si assemblano insieme in un anello a sei parti miste, un eteroesamero, che non può formarsi da nessuna delle due proteine da sola. Mediante crio-microscopia elettronica, il gruppo ha mostrato che questi eteroesameri sono costituiti da quattro unità di Hcp2 e due di Hcp3 disposte in un anello simmetrico, creando un poro rigido che può interagire con Bte1 ma non si mescola con gli anelli di Hcp1.
Un carico che cambia forma e un incastro a chiave e serratura
Strutture ad alta risoluzione dell'anello Hcp2–Hcp3 legato a Bte1 hanno rivelato che l'effettore deve riorganizzare in modo drastico parti della sua conformazione per adattarsi perfettamente al canale centrale. Diverse delle sue α-eliche oscillano di decine di gradi per evitare scontri con il tubo proteico, dimostrando quella che gli autori definiscono plasticità conformazionale. Quando hanno ingegnerizzato un legame disolfuro per bloccare una di queste eliche in posizione, Bte1 non è più stato in grado di legarsi efficacemente all'anello e la sua attività letale è diminuita nelle prove di competizione. La mappatura dettagliata delle superfici di contatto ha individuato aminoacidi specifici sia in Hcp2 che in Hcp3 essenziali per il legame e la secrezione. La mutazione di una singola residuo in uno dei partner è stata sufficiente a interrompere la consegna senza rompere l'anello stesso.
Anelli e tossine che coevolvono tra i lignaggi intestinali
Il gruppo si è poi chiesto se questo meccanismo sia unico per ceppi isolati o condiviso tra i lignaggi di B. fragilis. Confrontando coppie Hcp2–Hcp3 e i relativi geni degli effettori vicini in dozzine di ceppi, hanno scoperto che ogni lignaggio reca un effettore caratteristico in una regione variabile del locus e una coppia Hcp2–Hcp3 corrispondente. Scambi incrociati di queste coppie tra ceppi hanno mostrato che ogni anello misto riconosce solo il proprio effettore, nonostante anelli diversi possano comunque assemblarsi a livello strutturale. Hcp3, in particolare, contiene patch altamente variabili sulla superficie interna dell'anello che fungono da determinante principale di quale effettore possa legarsi. Nonostante questa diversità di sequenza, gli effettori condividono un dominio N-terminale conservato che si incastra nel ring come un modulo riutilizzabile, mentre i loro domini tossici C-terminali differiscono ampiamente.
Perché questo sistema a incastro conta
Nel complesso, questi risultati delineano un modello coevolutivo «a serratura e chiave»: la superficie interna dell'anello Hcp2–Hcp3 fornisce una serratura specifica per lignaggio, e il modulo N-terminale conservato di ciascun effettore funge da chiave che è stata sottilmente rimodellata in ceppi diversi. Le proteine effettrici possono flettersi e riorientarsi quanto basta per attraversare l'anello senza inceppare il tubo, consentendo la consegna di tossine relativamente grandi o complesse attraverso un canale stretto. Questo lavoro spiega come i simbionti intestinali regolino finemente le loro microlance molecolari per impiegare tossine distinte contro i rivali microbici, e suggerisce che riprogrammando queste interfacce a serratura e chiave i ricercatori potrebbero un giorno ingegnerizzare i sistemi di Tipo VI come piattaforme di consegna programmabili di molecole benefiche nelle comunità microbiche.
Citazione: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Parole chiave: sistema di secrezione di tipo VI, Bacteroides fragilis, competizione batterica, plasticità conformazionale delle proteine, ingegneria del microbioma