Clear Sky Science · ru
Конформационная пластичность эффектора обеспечивает родоспецифическую секрецию через гетерогексамеры Hcp у симбионтов кишечника
Микробные поединки внутри человеческого кишечника
В наших кишечниках обитают плотные сообщества микроорганизмов, которые постоянно соперничают за пространство и пищу. Чтобы выжить, некоторые бактерии используют молекулярное оружие, способное прокалывать и отравлять конкурентов, тем самым тонко формируя состав видов, преобладающих в кишке. В этом исследовании показано, как распространённый симбионт человека, Bacteroides fragilis, адаптирует крошечный инжекционный аппарат, чтобы доставлять специфические токсические белки в соседние клетки с поразительной точностью.
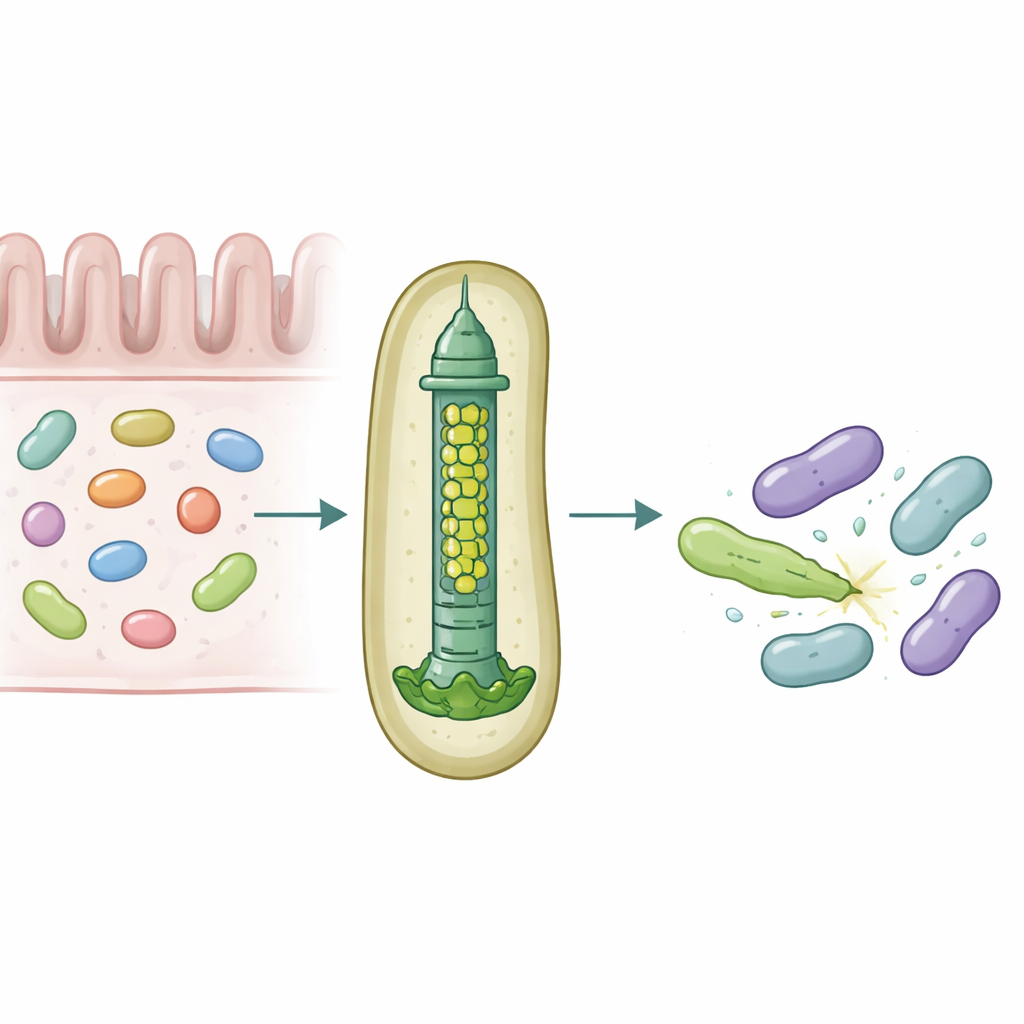
Микроскопический арбалет для бактериальной войны
Многие грамотрицательные бактерии используют аппарат, называемый системой секреции типа VI — пружинящее устройство, работающее как инвертированный хвост фага, которое стреляет токсическими «эффекторами» в близлежащие клетки. Центральную часть этой системы составляет труба, собранная из наслоенных колец белка Hcp. У большинства видов каждая такая система содержит одну форму Hcp, тогда как у B. fragilis, встречающегося в кишечнике человека, в одном локусе обнаружены пять различных генов hcp. Биологическая причина этой, казалось бы, избыточности была неизвестна, особенно учитывая необыкновенную эффективность этой системы в уничтожении близкородственных бактерий в лабораторных и животных моделях.
Специализированные кольца, собранные из смешанных частей
Исследователи объединили эксперименты по колонизации мышей, смешанные культуpы бактерий и биохимию белков, чтобы разобраться в ролях этих вариантов Hcp. Они обнаружили, что для эффективного убийства конкретным эффектором под названием Bte1 требуются только три — Hcp1, Hcp2 и Hcp3. Hcp1 образует основную трубу секрционной машины, как и ожидалось. Hcp2 и Hcp3, однако, выполняют другую функцию: они собираются вместе в смешанное шестичленное кольцо, гетерогексамер, которое не может сформироваться из одного только белка. С помощью крио-электронной микроскопии команда показала, что эти гетерогексамеры состоят из четырёх единиц Hcp2 и двух единиц Hcp3, расположенных в симметричном кольце, создающем жёсткое отверстие, способное взаимодействовать с Bte1, но не смешивающееся с кольцами Hcp1.
Изменяющий форму груз и механизм «замок и ключ»
Высокоразрешающие структуры кольца Hcp2–Hcp3 в комплексе с Bte1 показали, что эффектор вынужден существенно перестроить части своей формы, чтобы плотно разместиться в центральном канале. Несколько его α-спиралей отклоняются на десятки градусов, чтобы избежать столкновений с белковой трубой, демонстрируя то, что авторы называют конформационной пластичностью. Когда они создали дисульфидную связь, фиксирующую одну из этих спиралей, Bte1 уже не мог эффективно связываться с кольцом, и его активность в испытаниях на конкуренцию снизилась. Детальная картография контактных поверхностей указала конкретные аминокислоты на Hcp2 и Hcp3, необходимые для связывания и секреции. Мутация всего одной резидуи в любом из партнёров была достаточна, чтобы нарушить доставку, не разрушая при этом само кольцо.
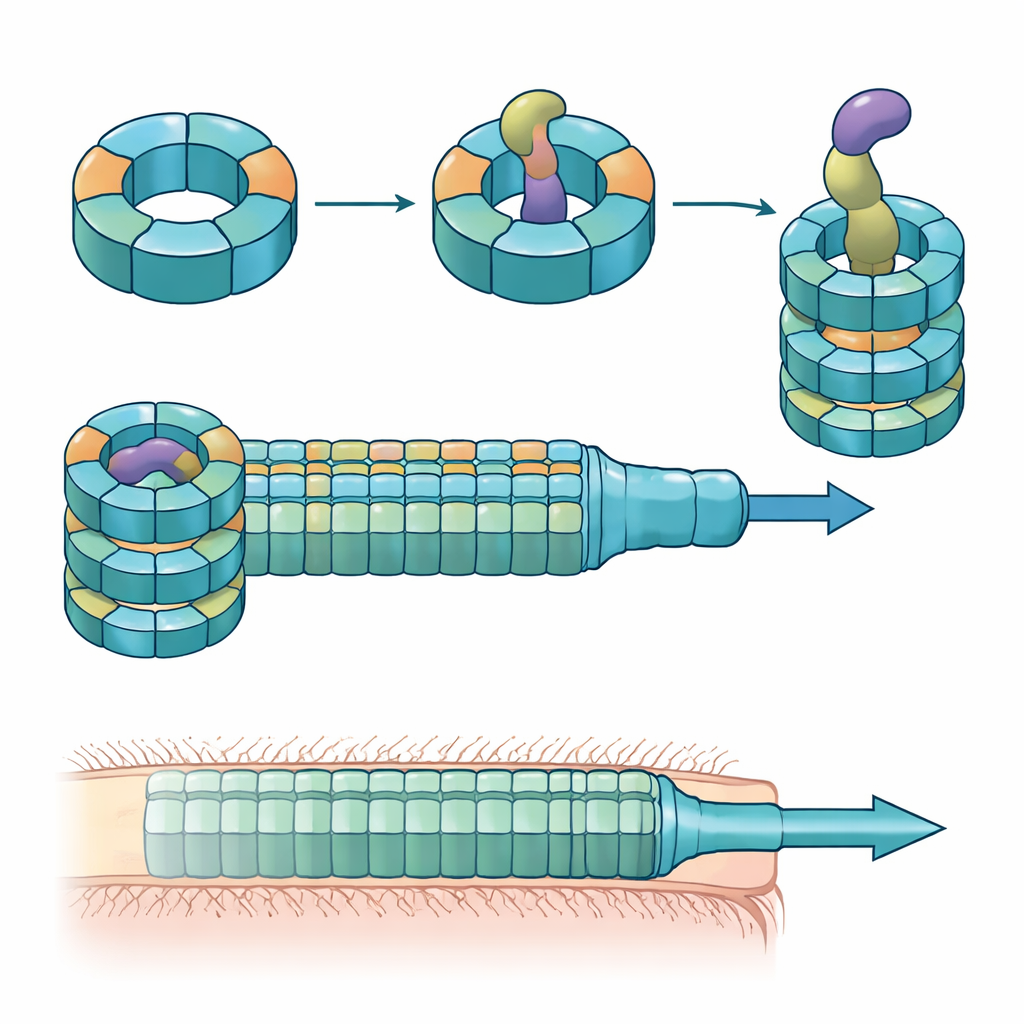
Коэволюция колец и токсинов в линиях кишечной микрофлоры
Затем команда задалась вопросом, уникален ли этот механизм для одной штамма или распространён среди линий B. fragilis. Сравнив пары Hcp2–Hcp3 и соседние гены эффекторных белков из десятков штаммов, они обнаружили, что каждая линия несёт характерный эффектор в переменной области локуса и соответствующую пару Hcp2–Hcp3. Перекрёстная перестановка этих пар между штаммами показала, что каждое смешанное кольцо распознаёт только свой собственный эффектор, хотя разные кольца по-прежнему могут собираться структурно. Hcp3, в частности, содержит сильно изменчивые участки на внутренней поверхности кольца, которые выступают главным детерминантом того, какой эффектор сможет связаться. Несмотря на эту последовательностную изменчивость, сами эффекторы имеют консервативную N-концевую складку, которая вставляется в кольцо как многоразовый модуль, в то время как их C-концевые токсические домены сильно различаются.
Почему эта система подгонки форм важна
Вместе эти результаты описывают модель коэволюции «замок и ключ»: внутренняя поверхность кольца Hcp2–Hcp3 служит родоспецифическим замком, а консервативный N-концевой модуль каждого эффектора — ключом, который в разных штаммах был слегка переработан. Эффекторные белки могут сгибаться и перестраиваться ровно настолько, чтобы протянуться через кольцо, не застревая в трубе, что позволяет доставлять относительно большие или сложные токсины через узкий канал. Эта работа объясняет, как симбионты кишечника тонко настраивают свои молекулярные гарпуны для применения разных токсинов против микробных конкурентов, и предполагает, что, перенастроив эти интерфейсы «замок‑и‑ключ», учёные однажды смогут сконструировать системы типа VI как программируемые платформы доставки полезных молекул в микробных сообществах.
Цитирование: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Ключевые слова: система секреции типа VI, Bacteroides fragilis, бактериальная конкуренция, конформационная пластичность белков, инжиниринг микробиома