Clear Sky Science · pt
A plasticidade conformacional do efetor possibilita secreção específica de linhagem via heterohexâmeros de Hcp em simbiontes intestinais
Duelo microbiano dentro do intestino humano
Nos nossos intestinos residem comunidades densas de microrganismos que constantemente disputam espaço e nutrientes. Para sobreviver, algumas bactérias empunham armas moleculares capazes de perfurar e envenenar rivais, influenciando de forma sutil quais espécies prosperam no intestino. Este estudo revela como um simbionte humano comum, Bacteroides fragilis, customiza um minúsculo aparelho de injeção para entregar proteínas tóxicas específicas a bactérias vizinhas com notável precisão.
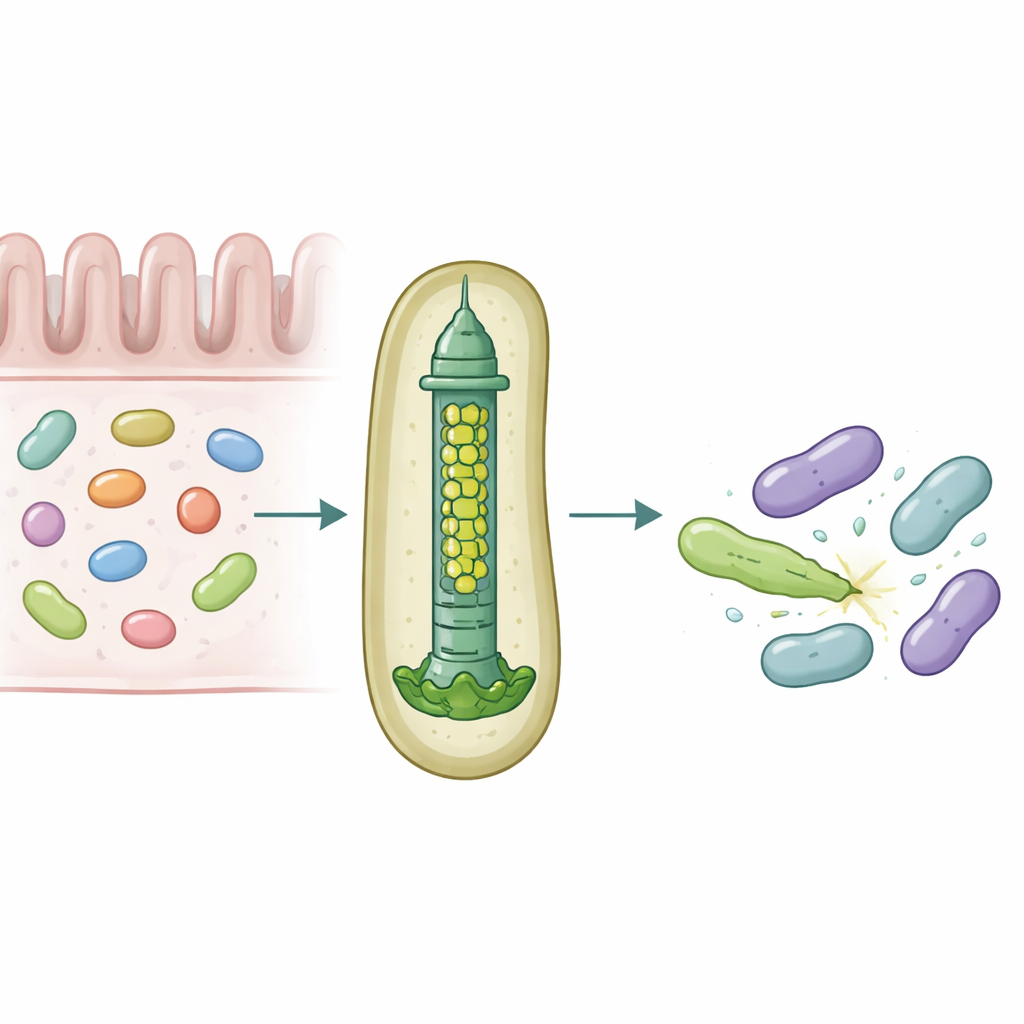
Uma lança microscópica para a guerra bacteriana
Muitas bactérias Gram-negativas usam um dispositivo chamado Sistema de Secreção Tipo VI, uma estrutura carregada por tensão que funciona como uma cauda de fago invertida, para disparar efetores tóxicos em células próximas. Uma parte central desse sistema é um tubo formado por anéis empilhados de uma proteína chamada Hcp. Na maioria das espécies, cada sistema possui uma única forma de Hcp, porém B. fragilis do intestino humano carrega cinco genes hcp diferentes em um único locus. A razão biológica para essa aparente redundância era desconhecida, principalmente porque esse sistema é incomumente potente ao matar bactérias intimamente relacionadas em modelos de laboratório e animais.
Anéis especializados construídos a partir de partes mistas
Os pesquisadores combinaram experimentos de colonização em camundongos, culturas bacterianas mistas e bioquímica de proteínas para dissecar as funções dessas variantes de Hcp. Eles descobriram que apenas três—Hcp1, Hcp2 e Hcp3—são necessários para a eliminação eficiente por um efetor específico chamado Bte1. Hcp1 forma o tubo principal da máquina de secreção, como esperado. Hcp2 e Hcp3, no entanto, fazem algo diferente: montam-se juntos em um anel de seis partes misto, um heterohexâmero, que não pode ser formado por nenhuma das proteínas isoladamente. Usando criomicroscopia eletrônica, a equipe mostrou que esses heterohexâmeros são compostos por quatro unidades de Hcp2 e duas de Hcp3 dispostas em um anel simétrico, criando um poro rígido que pode interagir com Bte1 mas não se mistura com os anéis de Hcp1.
Carga que muda de forma e um encaixe tipo fechadura-e-chave
Estruturas de alta resolução do anel Hcp2–Hcp3 ligado a Bte1 revelaram que o efetor precisa reorganizar dramaticamente partes de sua estrutura para se ajustar com firmeza dentro do canal central. Várias de suas α-hélices giram dezenas de graus para evitar colisões com o tubo proteico, demonstrando o que os autores chamam de plasticidade conformacional. Quando eles projetaram uma ligação dissulfeto para travar uma dessas hélices no lugar, Bte1 não conseguiu mais se ligar eficientemente ao anel, e sua atividade letal caiu em ensaios de competição. O mapeamento detalhado das superfícies de contato identificou aminoácidos específicos tanto em Hcp2 quanto em Hcp3 que são essenciais para a ligação e secreção. Mutar apenas um resíduo em qualquer um dos parceiros foi suficiente para interromper a entrega sem romper o anel em si.
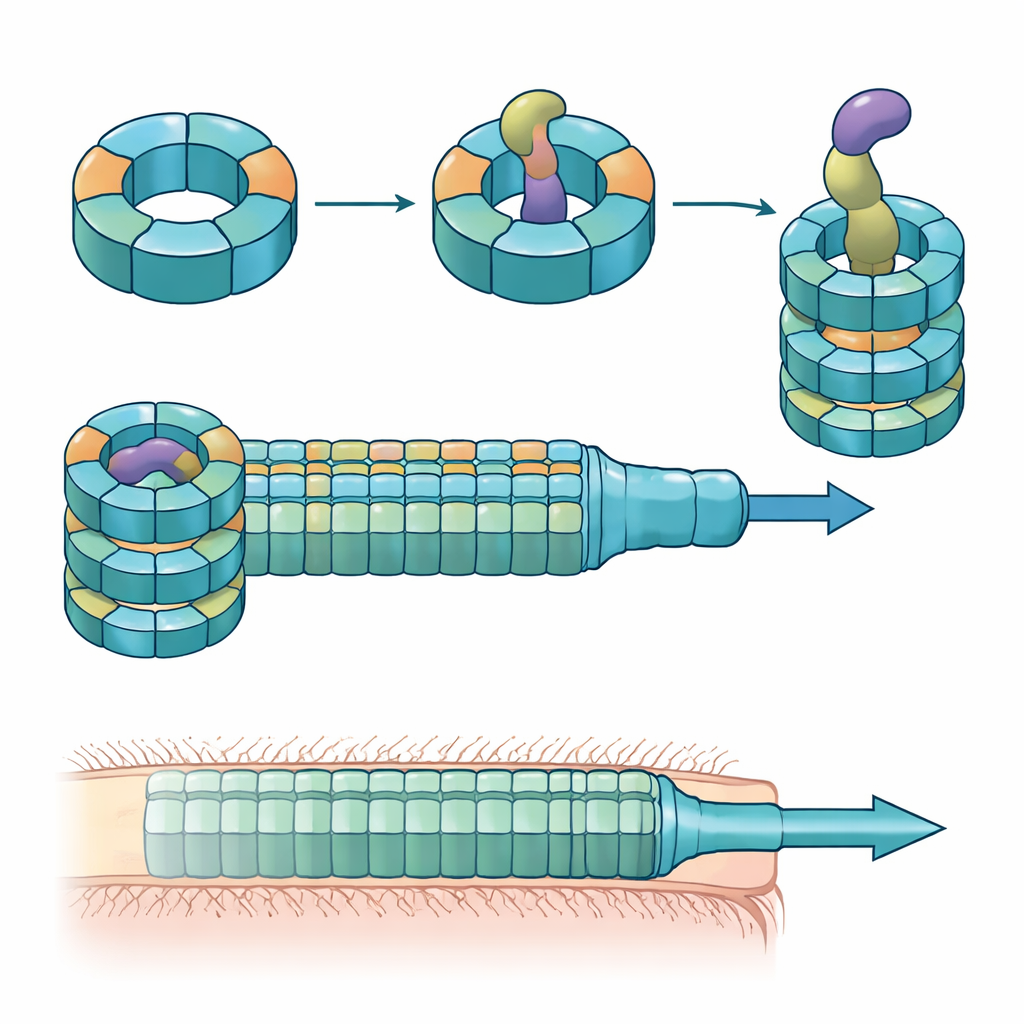
Anéis e toxinas coevoluindo entre linhagens intestinais
A equipe então investigou se esse mecanismo é exclusivo de uma linhagem ou compartilhado entre linhagens de B. fragilis. Ao comparar pares Hcp2–Hcp3 e seus genes efetores próximos em dezenas de cepas, eles descobriram que cada linhagem carrega um efetor característico em uma região variável do locus e um par correspondente Hcp2–Hcp3. A troca entre cepas desses pares mostrou que cada anel misto reconhece apenas seu próprio efetor, embora diferentes anéis ainda possam montar-se estruturalmente. Hcp3, em particular, contém áreas altamente variáveis na superfície interna do anel que atuam como o principal determinante de qual efetor pode se ligar. Apesar dessa diversidade de sequência, os efetores compartilham um domínio N-terminal conservado que se encaixa no anel como um módulo reutilizável, enquanto seus domínios C-terminais tóxicos variam amplamente.
Por que esse sistema de correspondência de formas é importante
Conjuntamente, esses achados delineiam um modelo coevolutivo do tipo "fechadura-e-chave": a superfície interna do anel Hcp2–Hcp3 fornece uma fechadura específica de linhagem, e o módulo N-terminal conservado de cada efetor funciona como uma chave que foi sutilmente remodelada em diferentes cepas. As proteínas efetoras podem flexionar e refoldar o suficiente para atravessar o anel sem travar o tubo, permitindo que toxinas relativamente grandes ou complexas sejam entregues através de um canal estreito. Este trabalho explica como simbiontes intestinais ajustam com precisão suas lanças moleculares para implantar toxinas distintas contra rivais microbianos, e sugere que, ao reconfigurar essas interfaces de fechadura-e-chave, pesquisadores poderão um dia projetar sistemas Tipo VI como plataformas de entrega programáveis para moléculas benéficas em comunidades microbianas.
Citação: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Palavras-chave: sistema de secreção tipo VI, Bacteroides fragilis, competição bacteriana, plasticidade conformacional de proteínas, engenharia do microbioma