Clear Sky Science · pl
Plastyczność konformacyjna efektora umożliwia wydzielanie specyficzne dla linii poprzez heteroheksamery Hcp u symbiontów jelitowych
Mikrobowe pojedynki w ludzkim jelicie
W naszych jelitach bytują gęste społeczności mikroorganizmów, które nieustannie rywalizują o przestrzeń i pokarm. Aby przetrwać, niektóre bakterie używają molekularnych broni mogących przebić i otruć rywali, subtelnie wpływając na to, które gatunki dominują w mikrobiocie. Badanie to ujawnia, jak powszechny symbiont jelitowy Bacteroides fragilis dostosowuje niewielki aparat wstrzykujący, aby z niezwykłą precyzją dostarczać specyficzne toksyczne białka do sąsiednich bakterii.
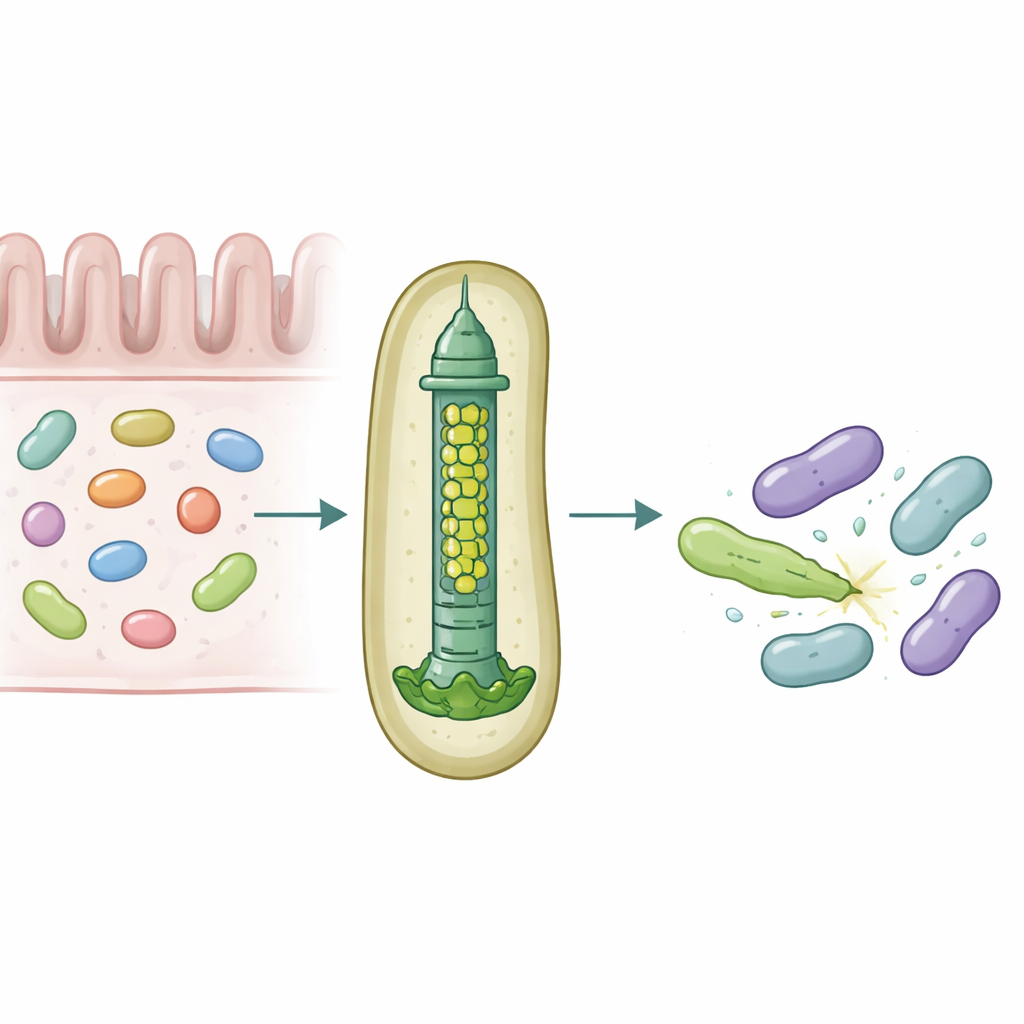
Microskopowy wyrzutnia włóczni do walki bakteryjnej
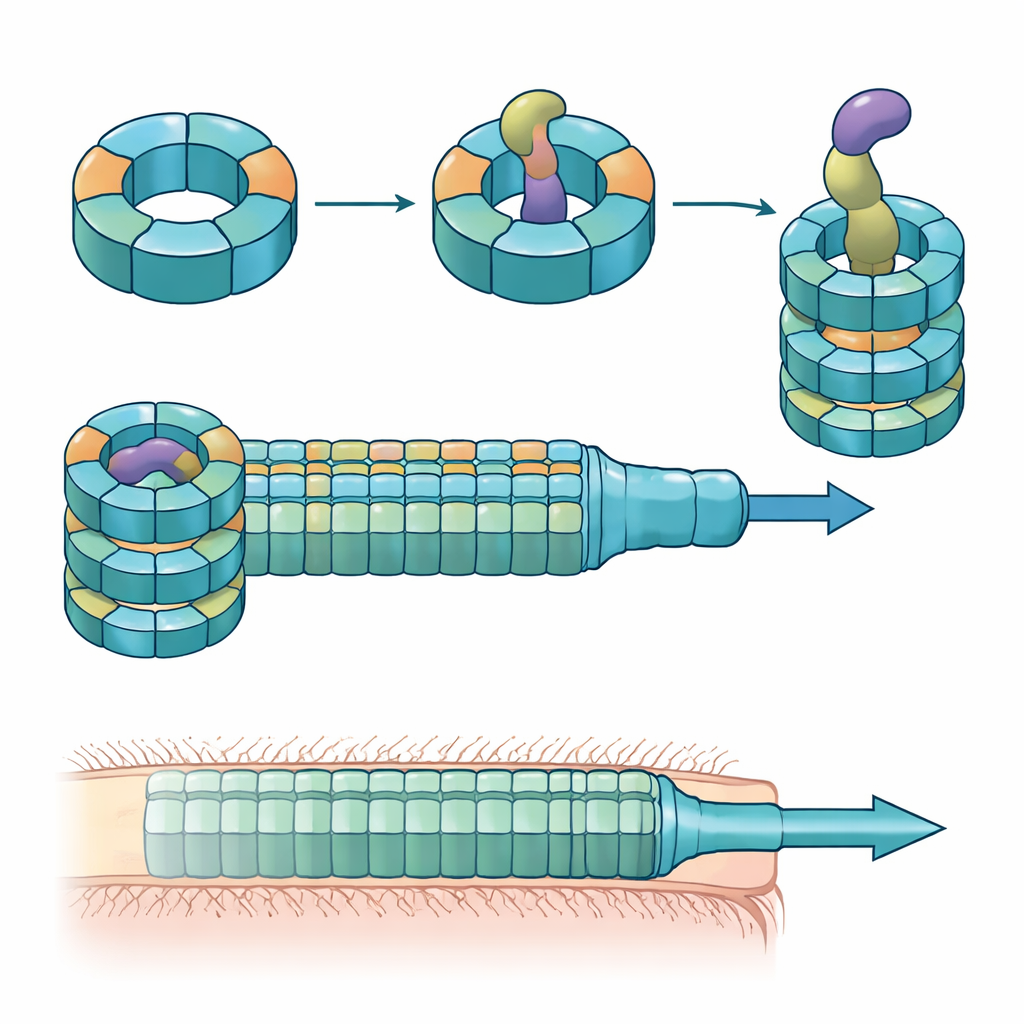
Wiele bakterii Gram-ujemnych używa urządzenia zwanego systemem sekrecji typu VI — sprężynującej struktury działającej jak odwrócony ogon fagowy — do wystrzeliwania toksycznych „efektorów” do pobliskich komórek. Centralną częścią tego systemu jest rurka zbudowana ze stosowanych pierścieni białka nazywanego Hcp. W większości gatunków każdy system ma jedną formę Hcp, tymczasem B. fragilis z ludzkiego jelita posiada pięć różnych genów hcp w pojedynczym locus. Biologicznego sensu tej pozornej redundancji nie znano, zwłaszcza że u tego gatunku system ten wyjątkowo skutecznie zabija blisko spokrewnione bakterie w modelach laboratoryjnych i zwierzęcych.
Specjalizowane pierścienie zbudowane z mieszanych części
Naukowcy połączyli eksperymenty kolonizacji myszy, hodowle mieszane bakterii i biochemię białek, aby rozpoznać role wariantów Hcp. Stwierdzili, że tylko trzy — Hcp1, Hcp2 i Hcp3 — są niezbędne do wydajnego zabijania przez konkretny efektor zwany Bte1. Hcp1 tworzy główną rurkę maszyny sekrecyjnej, jak można było przypuszczać. Hcp2 i Hcp3 robią jednak coś innego: składają się razem w wymieszany sześcioczłonowy pierścień, heteroheksamery, który nie może powstać z żadnego z tych białek osobno. Przy użyciu krio‑mikroskopii elektronowej zespół wykazał, że te heteroheksamery składają się z czterech jednostek Hcp2 i dwóch jednostek Hcp3 ułożonych w symetryczny pierścień, tworząc sztywny kanał, który potrafi zaangażować Bte1, ale nie łączy się z pierścieniami Hcp1.
Przekształcające się ładunki i dopasowanie typu zamek‑klucz
Struktury o wysokiej rozdzielczości pierścienia Hcp2–Hcp3 związane z Bte1 ujawniły, że efektor musi drastycznie przeorganizować fragmenty swojej struktury, by ciasno zmieścić się w centralnym kanale. Kilka jego α‑helis obraca się o dziesiątki stopni, aby uniknąć kolizji z rurką białkową, co autorzy określają jako plastyczność konformacyjną. Gdy zaprojektowali mostek disiarczkowy blokujący jedną z tych helis, Bte1 nie mógł już skutecznie wiązać się z pierścieniem, a jego aktywność zabójcza spadła w testach konkurencji. Szczegółowe mapowanie powierzchni kontaktowych wskazało konkretne aminokwasy zarówno na Hcp2, jak i Hcp3 niezbędne do wiązania i sekrecji. Zmutowanie zaledwie jednego resztu w dowolnym z partnerów wystarczyło, by zakłócić dostarczanie bez rozbijania samego pierścienia.
Współewoluujące pierścienie i toksyny w liniach jelitowych
Zespół zapytał następnie, czy ten mechanizm jest unikalny dla jednego szczepu, czy też występuje w różnych liniach B. fragilis. Porównując pary Hcp2–Hcp3 i ich pobliskie geny efektorowe z dziesiątek szczepów, odkryli, że każda linia niesie charakterystyczny efektor w zmiennej części locus i dopasowaną parę Hcp2–Hcp3. Wymiana tych par między szczepami pokazała, że każdy mieszany pierścień rozpoznaje tylko własny efektor, mimo że różne pierścienie potrafią nadal składać się strukturalnie. Hcp3 zawiera w szczególności silnie zmienne płaty na wewnętrznej powierzchni pierścienia, które działają jako główny wyznacznik, który efektor może się wiązać. Pomimo tej różnorodności sekwencji, efektory same w sobie dzielą zachowany N‑terminalny układ, który dokuje w pierścieniu jak moduł wielokrotnego użytku, podczas gdy ich toksyczne domeny C‑terminalne różnią się znacznie.
Dlaczego system dopasowania kształtów ma znaczenie
Razem te ustalenia opisują ewolucyjny model „zamek‑i‑klucz”: wewnętrzna powierzchnia pierścienia Hcp2–Hcp3 stanowi specyficzny dla linii zamek, a zachowany N‑terminalny moduł każdego efektora służy jako klucz, który w różnych szczepach został subtelnie przekształcony. Białka efektorowe potrafią zginać się i przefałdowywać na tyle, by przeciągnąć się przez pierścień bez zablokowania rurki, umożliwiając dostarczanie stosunkowo dużych lub złożonych toksyn przez wąski kanał. Praca ta wyjaśnia, jak symbionty jelitowe dopracowują swoje molekularne włócznie, aby wdrażać różne toksyny przeciwko mikrobiologicznym rywalom, i sugeruje, że przez przeprogramowanie tych interfejsów zamek‑i‑klucz badacze mogliby pewnego dnia zaprojektować systemy typu VI jako programowalne platformy dostarczające korzystne molekuły w społecznościach mikrobiologicznych.
Cytowanie: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Słowa kluczowe: system sekrecji typu VI, Bacteroides fragilis, konkurencja bakteryjna, plastyczność konformacyjna białek, inżynieria mikrobiomu