Clear Sky Science · nl
Conformationele plasticiteit van effectoren maakt lijnspecifieke secretie mogelijk via Hcp-heterohexameren in darm-symbionten
Microbiële duelletjes in de menselijke darm
Onze darmen herbergen dicht opeengepakte microbieele gemeenschappen die voortdurend om ruimte en voeding strijden. Om te overleven gebruiken sommige bacteriën moleculaire wapens die hun rivalen kunnen doorboren en vergiftigen, en zo subtiel bepalen welke soorten in de darm floreren. Deze studie onthult hoe een veelvoorkomende menselijke darm-symbiont, Bacteroides fragilis, een klein injectiemechanisme aanpast zodat het specifieke toxische eiwitten met opmerkelijke precisie in naburige bacteriën kan afleveren.
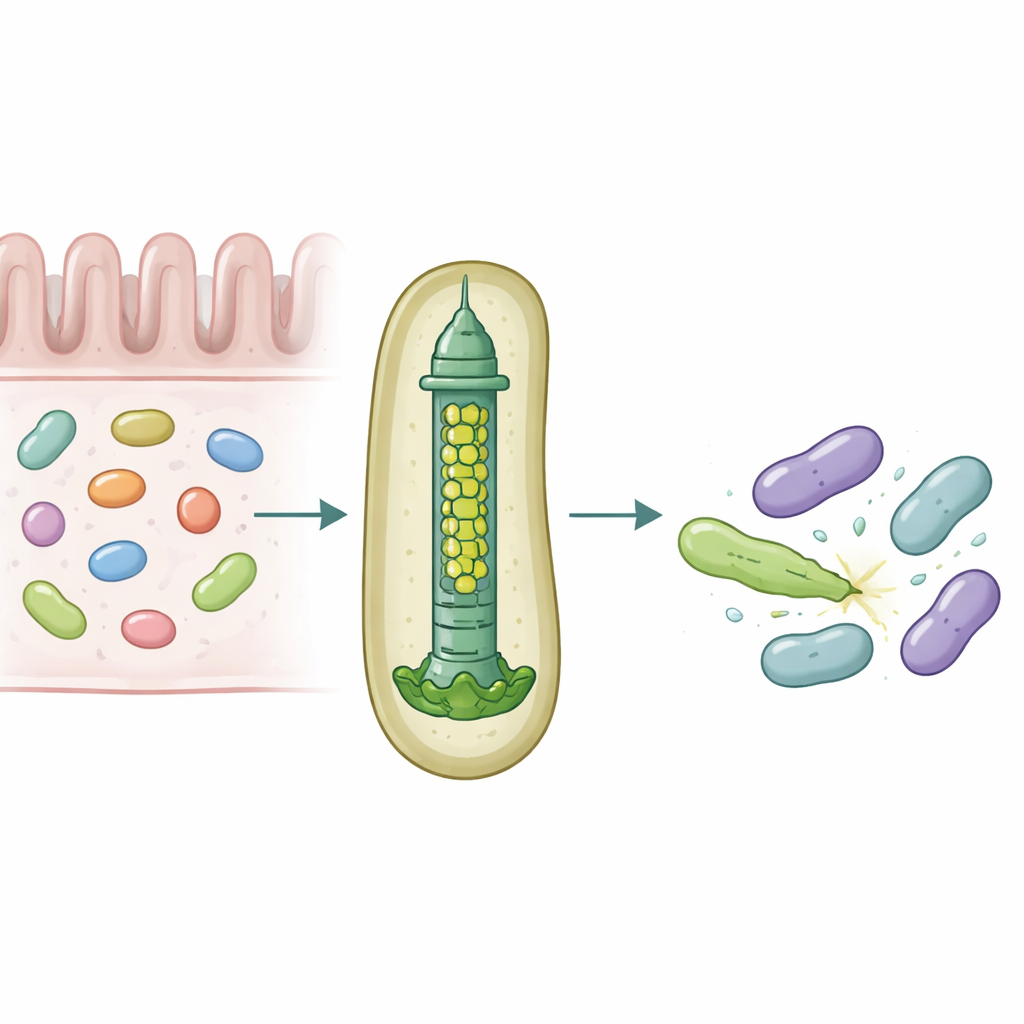
Een microscopisch speergeweer voor bacteriële oorlogsvoering
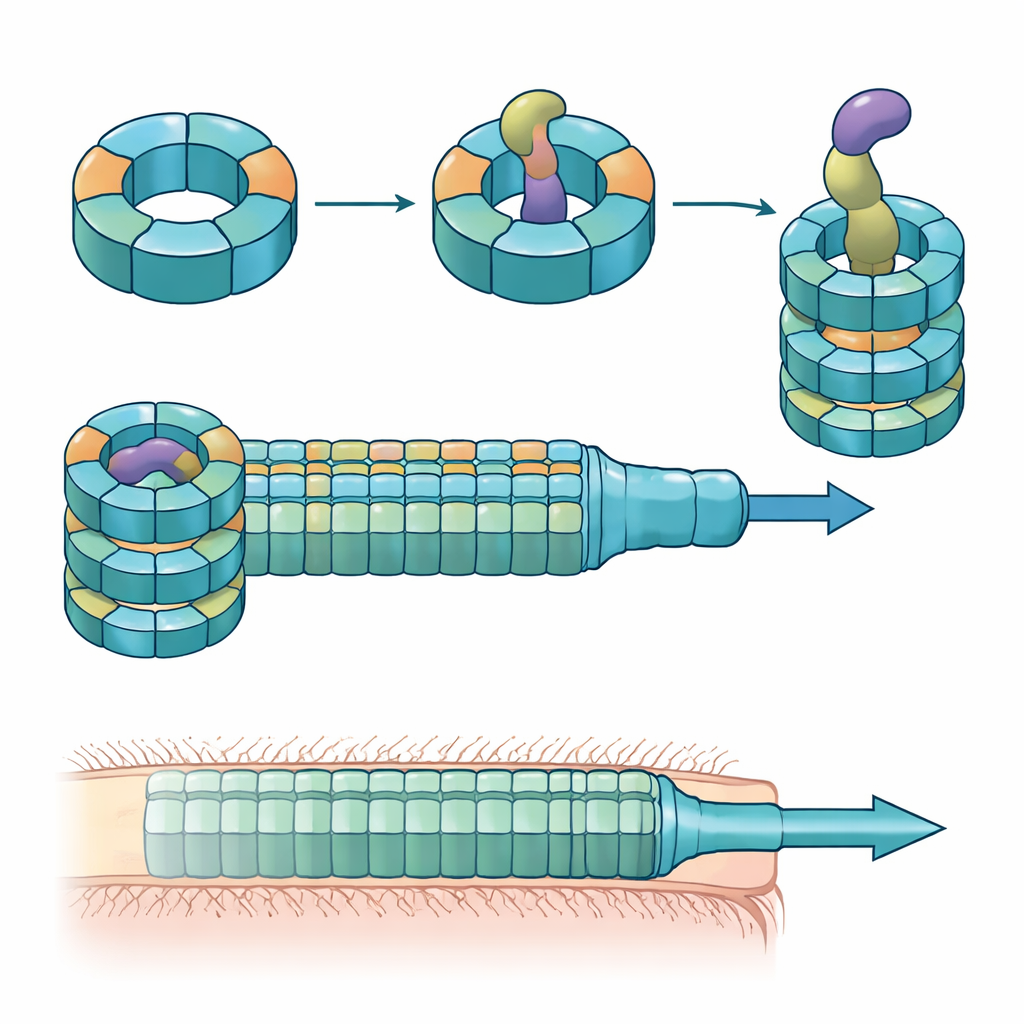
Veel Gram-negatieve bacteriën gebruiken een apparaat dat het Type VI-secretiesysteem wordt genoemd, een veerbelaste structuur die werkt als een omgekeerde fagestaart, om toxische "effectoren" in nabije cellen te schieten. Een centraal onderdeel van dit systeem is een buis opgebouwd uit opgestapelde ringen van een eiwit dat Hcp heet. Bij de meeste soorten heeft elk systeem één vorm van Hcp, maar B. fragilis uit de menselijke darm draagt vijf verschillende hcp-genen in één locus. De biologische reden voor deze schijnbare redundantie was onbekend, vooral omdat dit systeem ongewoon krachtig is in het doden van nauw verwante bacteriën in kweek en diermodellen.
Gespecialiseerde ringen gebouwd uit gemengde onderdelen
De onderzoekers combineerden muis-kolonisatie-experimenten, gemengde bacteriële kweeken en eiwitbiochemie om de rollen van deze Hcp-varianten te ontrafelen. Ze vonden dat slechts drie—Hcp1, Hcp2 en Hcp3—nodig zijn voor efficiënte doding door een specifieke effector genaamd Bte1. Hcp1 vormt zoals verwacht de hoofd-buis van het secretieapparaat. Hcp2 en Hcp3 doen echter iets anders: ze assembleren samen tot een gemengde zesdelige ring, een heterohexameer, die niet kan vormen uit één van beide eiwitten alleen. Met cryo-elektronenmicroscopie toonde het team aan dat deze heterohexameren bestaan uit vier Hcp2- en twee Hcp3-eenheden gerangschikt in een symmetrische ring, en zo een stijve porie creëren die Bte1 kan binden maar zich niet vermengt met Hcp1-ringen.
Vormveranderende lading en een slot-en-sleutelpassing
Hoge-resolutie structuren van de Hcp2–Hcp3-ring gebonden aan Bte1 onthulden dat de effector delen van zijn vorm ingrijpend moet herschikken om strak in het centrale kanaal te passen. Verschillende van zijn α-helices zwaaien tientallen graden om botsingen met de eiwitbuis te vermijden, wat de auteurs aanduiden als conformationele plasticiteit. Toen ze een disulfidebrug introduceerden om één van deze helices vast te zetten, kon Bte1 niet langer efficiënt aan de ring binden en daalde de dodekracht in competitie-assays. Gedetailleerde afbakening van de contactvlakken wees specifieke aminozuren op zowel Hcp2 als Hcp3 aan die essentieel zijn voor binding en secretie. Het muteren van slechts één residu op een van beide partners was genoeg om aflevering te verstoren zonder de ring zelf uiteen te laten vallen.
Co-evolverende ringen en toxines over darm-linages
Het team vroeg zich vervolgens af of dit mechanisme uniek is voor één stam of gedeeld wordt door meerdere B. fragilis-linages. Door Hcp2–Hcp3-paren en hun nabije effectorgenen van tientallen stammen te vergelijken, vonden ze dat elke linage een karakteristieke effector draagt in een variabele regio van de locus en een bijpassend Hcp2–Hcp3-paar. Het uitwisselen van deze paren tussen stammen toonde aan dat elke gemengde ring alleen zijn eigen effector herkent, ook al kunnen verschillende ringen structureel nog steeds assembleren. Hcp3 bevat in het bijzonder sterk variabele zones aan de binnenzijde van de ring die als de belangrijkste determinant fungeren voor welke effector kan binden. Ondanks deze sequentiediversiteit delen de effectoren zelf een geconserveerde N-terminale vouwing die in de ring wacht zoals een herbruikbare module, terwijl hun C-terminale toxische domeinen sterk variëren.
Waarom dit vorm-matching systeem van belang is
Gezamenlijk schetsen deze bevindingen een "slot-en-sleutel" co-evolutionair model: het binnenoppervlak van de Hcp2–Hcp3-ring biedt een linie-specifiek slot, en de geconserveerde N-terminale module van elke effector fungeert als een sleutel die in verschillende stammen subtiel is hervormd. Effector-eiwitten kunnen buigen en herschikken net genoeg om door de ring te schuiven zonder het kanaal te blokkeren, waardoor relatief grote of complexe toxines door een smalle doorgang geleverd kunnen worden. Dit werk verklaart hoe darm-symbionten hun moleculaire speerwapens verfijnen om verschillende toxines tegen microbiele rivalen in te zetten, en suggereert dat door het herschakelen van deze slot-en-sleutelinterfaces onderzoekers mogelijk Type VI-systemen in de toekomst kunnen ontwerpen als programmeerbare afleverplatforms voor nuttige moleculen in microbiële gemeenschappen.
Bronvermelding: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Trefwoorden: type VI-secretiesysteem, Bacteroides fragilis, bacteriële competitie, eiwitconformationele plasticiteit, microbioom-engineering