Clear Sky Science · fr
La plasticité conformationnelle des effecteurs permet une sécrétion spécifique à la lignée via des hétérohexamères Hcp chez les symbiotes intestinaux
Duels microbien dans l’intestin humain
Nos intestins abritent des communautés microbiennes denses qui se disputent en permanence l’espace et les ressources. Pour survivre, certaines bactéries utilisent des armes moléculaires capables de percer et d’empoisonner leurs rivales, façonnant discrètement les espèces qui prospèrent dans le microbiote. Cette étude révèle comment un symbiote intestinal humain courant, Bacteroides fragilis, adapte un minuscule appareil d’injection pour délivrer des protéines toxiques spécifiques à des bactéries voisines avec une précision remarquable.
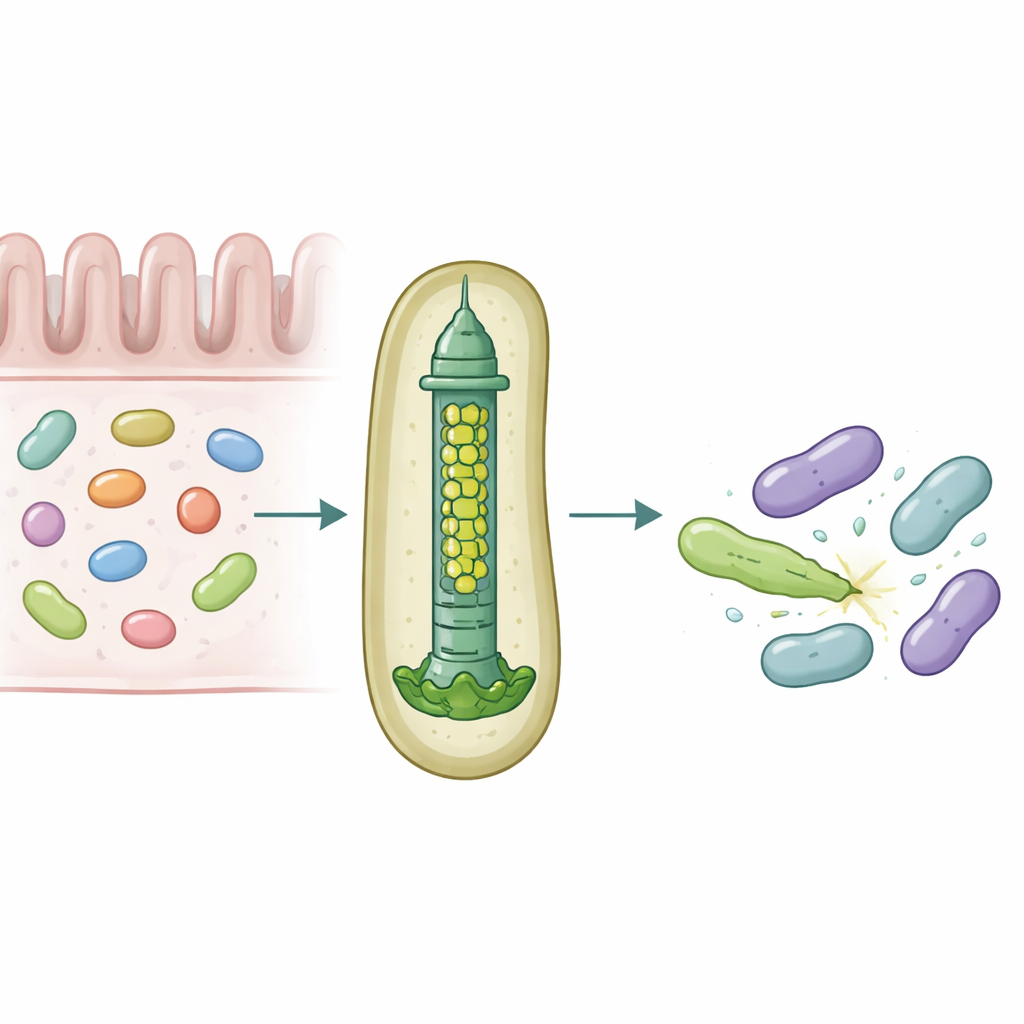
Une lance microscopique pour la guerre bactérienne
De nombreuses bactéries à Gram négatif emploient un dispositif nommé système de sécrétion de type VI, une structure à ressort qui fonctionne comme une queue de phage inversée, pour propulser des « effecteurs » toxiques dans les cellules voisines. Une partie centrale de ce système est un tube formé d’anneaux empilés d’une protéine appelée Hcp. Chez la plupart des espèces, chaque système possède une seule forme d’Hcp, pourtant B. fragilis issu de l’intestin humain porte cinq gènes hcp différents dans un même locus. La raison biologique de cette redondance apparente était inconnue, d’autant que ce système est exceptionnellement efficace pour tuer des bactéries étroitement apparentées dans des modèles de laboratoire et d’animal.
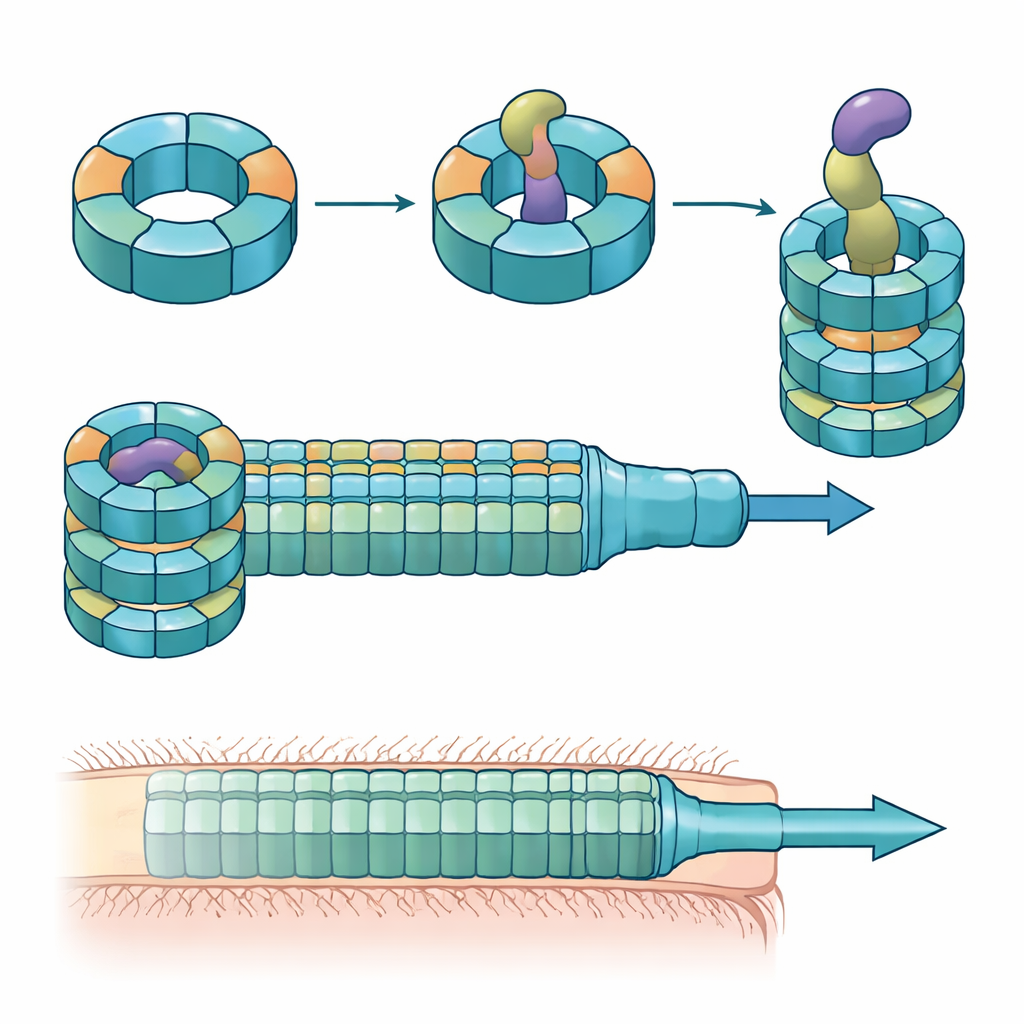
Anneaux spécialisés construits à partir de pièces mélangées
Les chercheurs ont combiné des expériences de colonisation chez la souris, des cultures bactériennes mixtes et de la biochimie protéique pour disséquer les rôles de ces variantes d’Hcp. Ils ont constaté que seuls trois — Hcp1, Hcp2 et Hcp3 — sont nécessaires pour une mise à mort efficace par un effecteur particulier appelé Bte1. Hcp1 forme le tube principal de la machine de sécrétion, comme prévu. Hcp2 et Hcp3, en revanche, font autre chose : ils s’assemblent ensemble en un anneau mixte à six éléments, un hétérohexamère, qui ne peut pas se former à partir de l’un ou l’autre protéine seul. Grâce à la cryo‑microscopie électronique, l’équipe a montré que ces hétérohexamères sont constitués de quatre unités Hcp2 et de deux unités Hcp3 disposées en anneau symétrique, créant un pore rigide capable d’accueillir Bte1 mais ne se mélangeant pas avec les anneaux Hcp1.
Cargaison changeante de forme et ajustement clé‑serrure
Les structures à haute résolution de l’anneau Hcp2–Hcp3 lié à Bte1 ont révélé que l’effecteur doit réarranger fortement des portions de sa conformation pour s’emboîter confortablement dans le canal central. Plusieurs de ses hélices α basculent de plusieurs dizaines de degrés pour éviter des collisions avec le tube protéique, illustrant ce que les auteurs appellent la plasticité conformationnelle. Lorsqu’ils ont conçu une liaison disulfure pour verrouiller l’une de ces hélices, Bte1 n’a plus pu se lier efficacement à l’anneau et son activité létale a chuté dans les tests de compétition. Une cartographie détaillée des surfaces de contact a identifié des acides aminés spécifiques sur Hcp2 et Hcp3 essentiels à la liaison et à la sécrétion. La mutation d’un seul résidu sur l’un ou l’autre partenaire suffisait à perturber la livraison sans rompre l’anneau lui‑même.
Anneaux et toxines co‑évoluant entre lignées intestinales
L’équipe s’est ensuite demandé si ce mécanisme est propre à une souche ou partagé entre les lignées de B. fragilis. En comparant les paires Hcp2–Hcp3 et leurs gènes effecteurs voisins dans des dizaines de souches, ils ont découvert que chaque lignée porte un effecteur caractéristique dans une région variable du locus et une paire Hcp2–Hcp3 assortie. L’échange croisé de ces paires entre souches a montré que chaque anneau mixte ne reconnaît que son propre effecteur, même si différents anneaux peuvent encore s’assembler structurellement. Hcp3, en particulier, contient des zones fortement variables sur la surface interne de l’anneau qui servent de déterminant principal pour le choix de l’effecteur pouvant se lier. Malgré cette diversité de séquence, les effecteurs partagent eux‑mêmes un repli N‑terminal conservé qui s’amarre dans l’anneau comme un module réutilisable, tandis que leurs domaines toxiques C‑terminaux diffèrent largement.
Pourquoi ce système d’ajustement forme‑contre‑forme est important
Ensemble, ces résultats décrivent un modèle co‑évolutif « clé‑serrure » : la surface interne de l’anneau Hcp2–Hcp3 fournit une serrure spécifique à la lignée, et le module N‑terminal conservé de chaque effecteur fait office de clé subtilement remodelée selon les souches. Les protéines effectrices peuvent se plier et se reconfigurer juste assez pour passer à travers l’anneau sans bloquer le tube, permettant la livraison de toxines relativement grandes ou complexes par un canal étroit. Ce travail explique comment les symbiotes intestinaux ajustent finement leurs lances moléculaires pour déployer des toxines distinctes contre des rivaux microbien, et suggère qu’en reprogrammant ces interfaces clé‑serrure, les chercheurs pourraient un jour concevoir des systèmes de type VI comme plateformes de livraison programmables de molécules bénéfiques dans les communautés microbiennes.
Citation: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Mots-clés: système de sécrétion de type VI, Bacteroides fragilis, compétition bactérienne, plasticité conformationnelle des protéines, ingénierie du microbiome