Clear Sky Science · es
La plasticidad conformacional de los efectores permite una secreción específica por linaje mediante heterohexámeros de Hcp en simbiontes intestinales
Duelo microbiano dentro del intestino humano
Nuestros intestinos albergan comunidades densas de microbios que compiten constantemente por espacio y nutrientes. Para sobrevivir, algunas bacterias emplean armas moleculares que pueden perforar y envenenar a sus rivales, moldeando de forma sutil qué especies prosperan en el intestino. Este estudio revela cómo un simbionte intestinal humano común, Bacteroides fragilis, personaliza un diminuto aparato de inyección para entregar proteínas tóxicas específicas a bacterias vecinas con notable precisión.
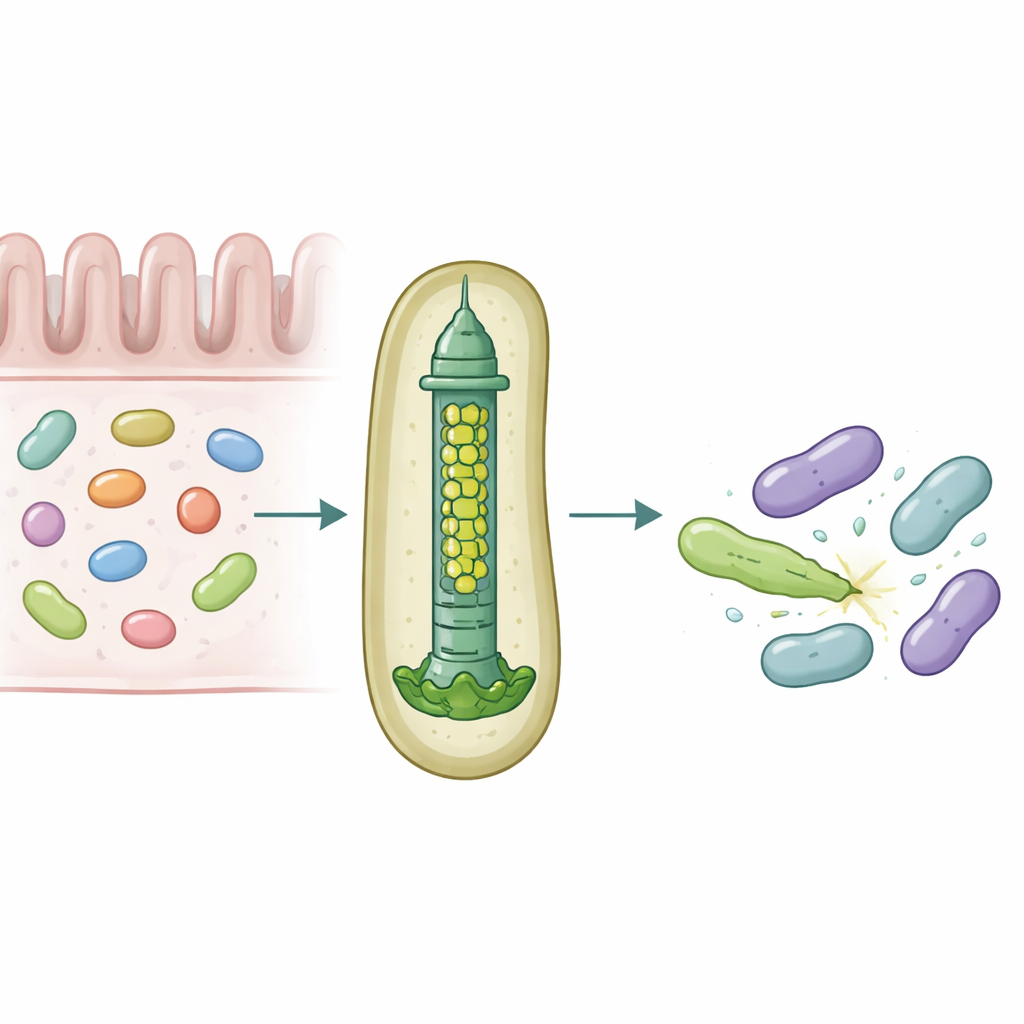
Una micrométrica ballesta para la guerra bacteriana
Muchas bacterias Gramnegativas usan un dispositivo llamado Sistema de Secreción Tipo VI, una estructura con resorte que funciona como una cola de fago invertida, para disparar «efectores» tóxicos a células cercanas. Una parte central de este sistema es un tubo formado por anillos apilados de una proteína llamada Hcp. En la mayoría de especies, cada sistema tiene una única forma de Hcp; sin embargo, B. fragilis del intestino humano porta cinco genes hcp diferentes en un único locus. La razón biológica de esta aparente redundancia era desconocida, sobre todo porque este sistema es inusualmente potente para matar bacterias estrechamente relacionadas en cultivos y modelos animales.
Anillos especializados construidos con piezas mixtas
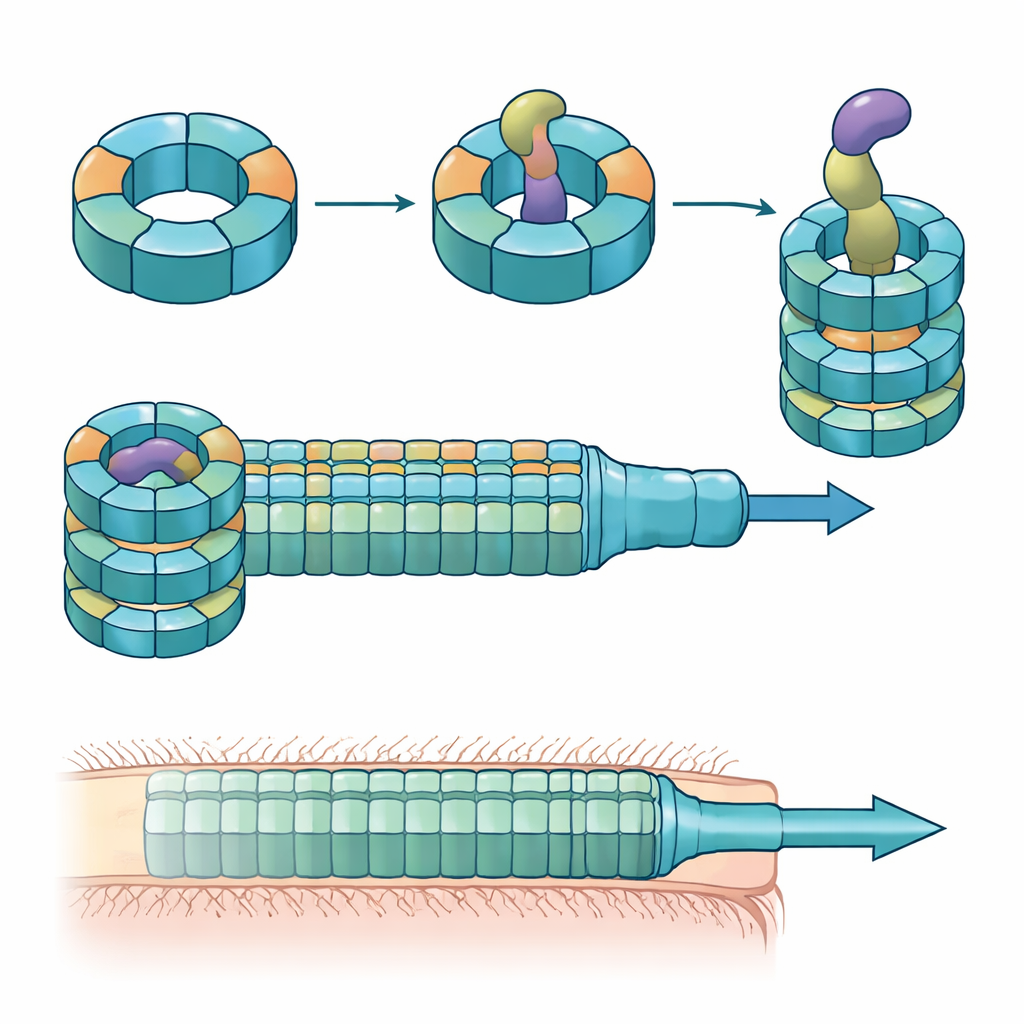
Los investigadores combinaron experimentos de colonización en ratón, cultivos bacterianos mixtos y bioquímica proteica para desentrañar los roles de estas variantes de Hcp. Encontraron que solo tres—Hcp1, Hcp2 y Hcp3—son necesarias para una eliminación eficiente por parte de un efector particular llamado Bte1. Hcp1 forma el tubo principal de la máquina de secreción, como se esperaba. Hcp2 y Hcp3, sin embargo, hacen algo distinto: se ensamblan juntos en un anillo de seis partes mixtas, un heterohexámero, que no puede formarse a partir de cualquiera de las proteínas por sí sola. Mediante crio-microscopía electrónica, el equipo mostró que estos heterohexámeros están compuestos por cuatro subunidades de Hcp2 y dos de Hcp3 dispuestas en un anillo simétrico, creando un poro rígido que puede interactuar con Bte1 pero que no se mezcla con los anillos de Hcp1.
Carga que cambia de forma y un ajuste de llave y cerradura
Estructuras de alta resolución del anillo Hcp2–Hcp3 unido a Bte1 revelaron que el efector debe reorganizar drásticamente partes de su conformación para encajar con firmeza dentro del canal central. Varias de sus hélices α se abatieron decenas de grados para evitar colisiones con el tubo proteico, demostrando lo que los autores denominan plasticidad conformacional. Cuando introdujeron un puente disulfuro para inmovilizar una de estas hélices, Bte1 ya no pudo unirse eficientemente al anillo y su actividad letal disminuyó en ensayos de competencia. El mapeo detallado de las superficies de contacto identificó aminoácidos concretos tanto en Hcp2 como en Hcp3 que son esenciales para la unión y la secreción. Mutar solo un residuo en cualquiera de las dos partes fue suficiente para interrumpir la entrega sin deshacer el anillo en sí.
Anillos y toxinas coevolucionando entre linajes intestinales
El equipo preguntó entonces si este mecanismo es único de una cepa o si se comparte entre linajes de B. fragilis. Al comparar pares Hcp2–Hcp3 y sus genes de efectores cercanos en docenas de cepas, hallaron que cada linaje porta un efector característico en una región variable del locus y un par Hcp2–Hcp3 a juego. El intercambio cruzado de estos pares entre cepas mostró que cada anillo mixto solo reconoce a su propio efector, aunque diferentes anillos todavía pueden ensamblarse estructuralmente. Hcp3, en particular, contiene parches altamente variables en la superficie interna del anillo que actúan como el determinante principal de qué efector puede unirse. A pesar de esta diversidad de secuencia, los efectores comparten un pliegue N-terminal conservado que se acopla en el anillo como un módulo reutilizable, mientras que sus dominios tóxicos C-terminales difieren ampliamente.
Por qué importa este sistema de coincidencia de formas
En conjunto, estos hallazgos delinean un modelo coevolutivo de «llave y cerradura»: la superficie interna del anillo Hcp2–Hcp3 proporciona una cerradura específica de linaje, y el módulo N-terminal conservado de cada efector actúa como una llave que ha sido sutilmente remodelada en diferentes cepas. Las proteínas efectoras pueden flexionarse y replegarse lo justo para atravesar el anillo sin atascar el tubo, permitiendo que toxinas relativamente grandes o complejas se entreguen a través de un canal estrecho. Este trabajo explica cómo los simbiontes intestinales ajustan sus micrométricas lanzas moleculares para desplegar toxinas distintas contra competidores microbianos, y sugiere que al reconfigurar estas interfaces de llave y cerradura, los investigadores podrían algún día diseñar sistemas Tipo VI como plataformas de entrega programables de moléculas beneficiosas en comunidades microbianas.
Cita: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Palabras clave: sistema de secreción tipo VI, Bacteroides fragilis, competencia bacteriana, plasticidad conformacional de proteínas, ingeniería del microbioma